

3',4'-Dihydroxyflavonol down-regulates monocyte chemoattractant protein-1 in smooth muscle: role of focal adhesion kinase and PDGF receptor signalling
- PMID: 19371329
- PMCID: PMC2707972
- DOI: 10.1111/j.1476-5381.2009.00199.x
3',4'-Dihydroxyflavonol down-regulates monocyte chemoattractant protein-1 in smooth muscle: role of focal adhesion kinase and PDGF receptor signalling
Abstract
Background and purpose: We investigated the effects of a synthetic flavonol, 3',4'-dihydroxyflavonol (DiOHF) on the expression of monocyte chemoattractant protein-1 (MCP-1) in rat vascular smooth muscle cells.
Experimental approach: MCP-1 expression was assessed by quantitative real-time PCR and protein phosphorylation by immunoprecipitation and Western blots.
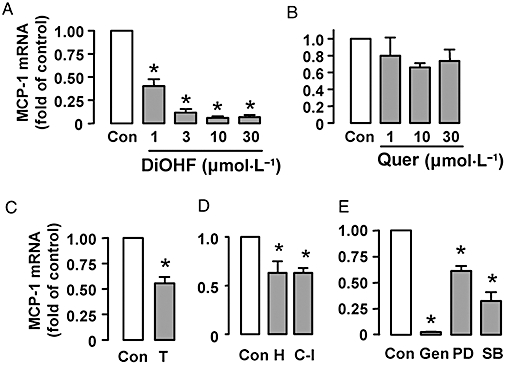
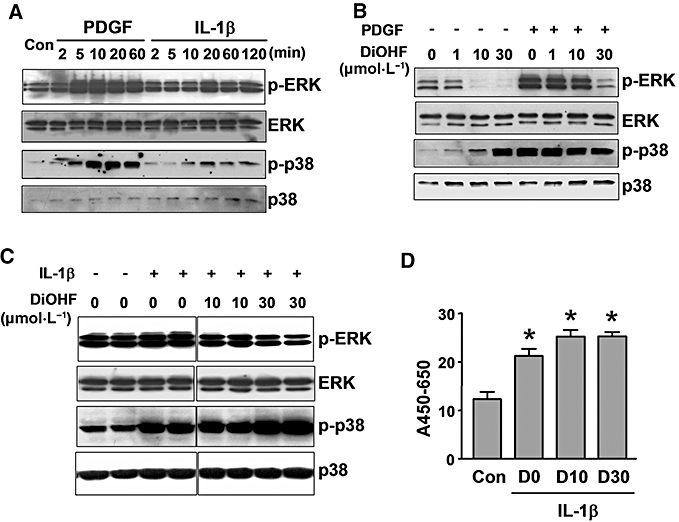
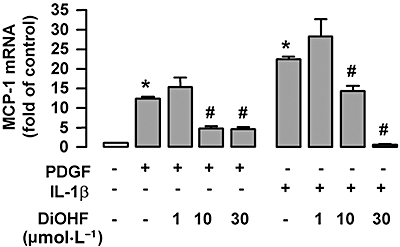
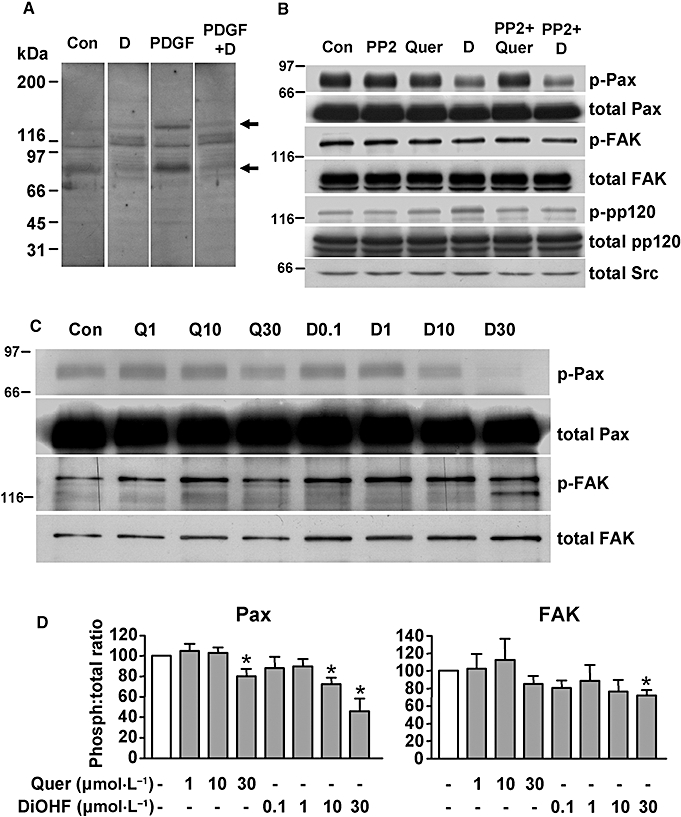
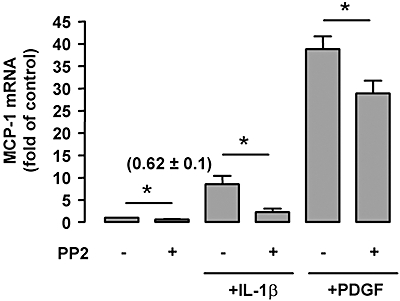
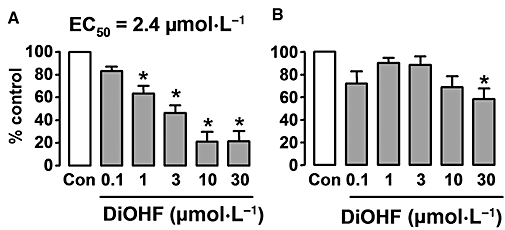
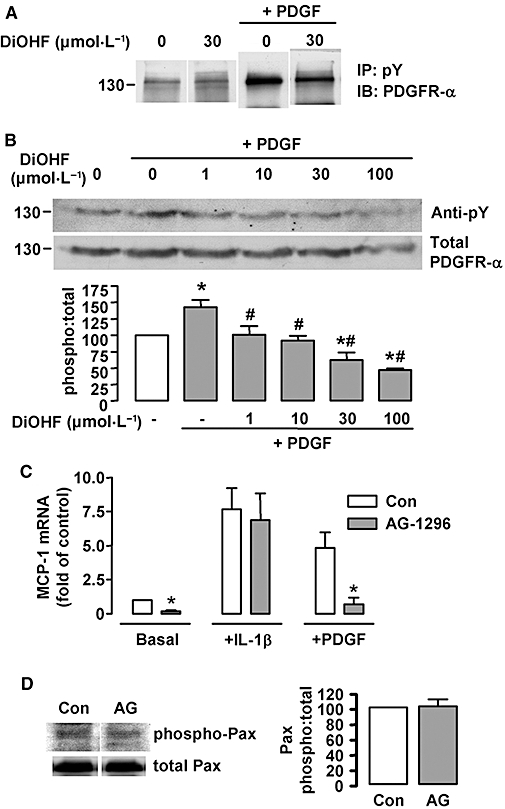
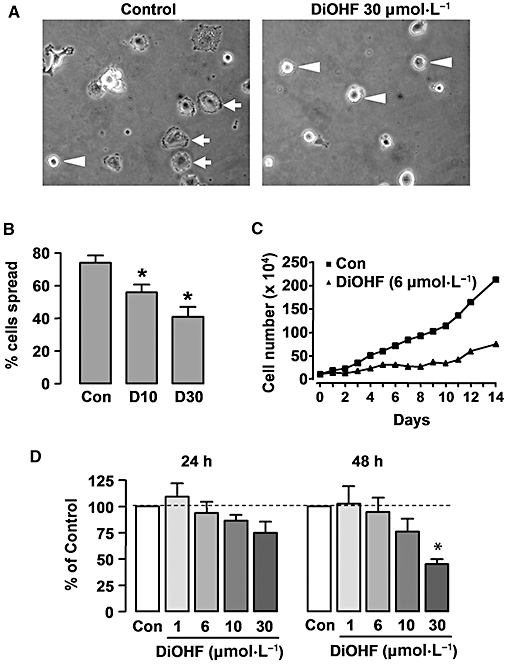
Key results: DiOHF (1-30 micromol x L(-1)) concentration-dependently reduced MCP-1 expression in both quiescent cells and cells stimulated with platelet-derived growth factor (PDGF) or interleukin 1-beta. The effect of DiOHF was associated with a suppression of focal adhesion kinase (FAK)-mediated signalling. In vitro kinase assays demonstrated that DiOHF is a potent inhibitor of FAK kinase activity (EC(50)= 2.4 micromol x L(-1)). Expression of FAK-related non-kinase reduced basal MCP-1 expression, but not that induced by PDGF or interleukin 1-beta. DiOHF also inhibited autophosphorylation of PDGF receptors. The PDGF receptor inhibitor AG-1296 potently suppressed basal and PDGF-induced MCP-1 expression. Inhibition of extracellular signal-regulated kinase activation by DiOHF, either directly or indirectly, may also be involved in its effects on MCP-1 expression. DiOHF had no inhibitory effect on either p38 or nuclear factor-kappaB activation. Moreover, DiOHF inhibited smooth muscle cell spreading (a FAK-mediated response) and proliferation.
Conclusions and implications: This is the first report on a flavonoid compound (DiOHF) that is a potent FAK inhibitor. DiOHF also inhibits PDGF receptor autophosphorylation. These effects underlie the inhibitory action of DiOHF on MCP-1 expression in smooth muscle cells. Our results suggest that DiOHF might be a useful tool for dissection of the (patho)physiological roles of FAK signalling.
Figures

Similar articles
-
Platelet-derived growth factor-BB (PDGF-BB) regulation of migration and focal adhesion kinase phosphorylation in rabbit aortic vascular smooth muscle cells: roles of phosphatidylinositol 3-kinase and mitogen-activated protein kinases.Cardiovasc Res. 1999 Mar;41(3):708-21. doi: 10.1016/s0008-6363(98)00232-6. Cardiovasc Res. 1999. PMID: 10435043
-
Focal adhesion kinase facilitates platelet-derived growth factor-BB-stimulated ERK2 activation required for chemotaxis migration of vascular smooth muscle cells.J Biol Chem. 2000 Dec 29;275(52):41092-9. doi: 10.1074/jbc.M005450200. J Biol Chem. 2000. PMID: 10998418
-
Role of JNK, p38, and ERK in platelet-derived growth factor-induced vascular proliferation, migration, and gene expression.Arterioscler Thromb Vasc Biol. 2003 May 1;23(5):795-801. doi: 10.1161/01.ATV.0000066132.32063.F2. Epub 2003 Mar 13. Arterioscler Thromb Vasc Biol. 2003. PMID: 12637337
-
Anthocyanidins, novel FAK inhibitors, attenuate PDGF-BB-induced aortic smooth muscle cell migration and neointima formation.Cardiovasc Res. 2014 Mar 1;101(3):503-12. doi: 10.1093/cvr/cvt337. Epub 2013 Dec 20. Cardiovasc Res. 2014. PMID: 24363205
-
Synergistic induction of monocyte chemoattractant protein-1 by integrins and platelet-derived growth factor via focal adhesion kinase in mesangial cells.Nephrol Dial Transplant. 2005 Oct;20(10):2080-8. doi: 10.1093/ndt/gfh998. Epub 2005 Jul 19. Nephrol Dial Transplant. 2005. PMID: 16030037
Cited by
-
TRAIL/DR5 signaling promotes macrophage foam cell formation by modulating scavenger receptor expression.PLoS One. 2014 Jan 22;9(1):e87059. doi: 10.1371/journal.pone.0087059. eCollection 2014. PLoS One. 2014. PMID: 24466325 Free PMC article.
-
Macrophage-stimulated microRNA expression in mural cells promotes transplantation-induced neointima formation.Oncotarget. 2017 May 2;8(18):30100-30111. doi: 10.18632/oncotarget.16279. Oncotarget. 2017. PMID: 28415796 Free PMC article.
-
Flavonols in the Prevention of Diabetes-induced Vascular Dysfunction.J Cardiovasc Pharmacol. 2015 Jun;65(6):532-44. doi: 10.1097/FJC.0000000000000180. J Cardiovasc Pharmacol. 2015. PMID: 25387248 Free PMC article. Review.
-
The antioxidant compound tert-butylhydroquinone activates Akt in myocardium, suppresses apoptosis and ameliorates pressure overload-induced cardiac dysfunction.Sci Rep. 2015 Aug 11;5:13005. doi: 10.1038/srep13005. Sci Rep. 2015. PMID: 26260024 Free PMC article.
References
-
- Aiello RJ, Bourassa PA, Lindsey S, Weng W, Natoli E, Rollins BJ, et al. Monocyte chemoattractant protein-1 accelerates atherosclerosis in apolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 1999;19:1518–1525. - PubMed
-
- Birdsall HH, Green DM, Trial J, Youker KA, Burns AR, MacKay CR, et al. Complement C5a, TGF-beta 1, and MCP-1, in sequence, induce migration of monocytes into ischemic canine myocardium within the first one to five hours after reperfusion. Circulation. 1997;95:684–692. - PubMed
-
- Cartel NJ, Liu J, Wang J, Post M. PDGF-BB-mediated activation of p42(MAPK) is independent of PDGF beta-receptor tyrosine phosphorylation. Am J Physiol Lung Cell Mol Physiol. 2001;281:L786–L798. - PubMed
-
- Charo IF, Taubman MB. Chemokines in the pathogenesis of vascular disease. Circ Res. 2004;95:858–866. - PubMed
-
- Chen Y, Hallenbeck JM, Ruetzler C, Bol D, Thomas K, Berman NE, et al. Overexpression of monocyte chemoattractant protein 1 in the brain exacerbates ischemic brain injury and is associated with recruitment of inflammatory cells. J Cereb Blood Flow Metab. 2003;23:748–755. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
