

In vivo detection of residues required for ligand-selective activation of the S-locus receptor in Arabidopsis
- PMID: 19375322
- PMCID: PMC2747293
- DOI: 10.1016/j.cub.2009.03.037
In vivo detection of residues required for ligand-selective activation of the S-locus receptor in Arabidopsis
Abstract
The self-incompatibility response of crucifers is a barrier to fertilization in which arrest of pollen tube development is mediated by allele-specific interactions between polymorphic receptors and ligands encoded by the S-locus haplotype. Activation of stigma-expressed S-locus receptor kinase (SRK) [1] by pollen coat-localized S-locus cysteine-rich (SCR) ligand [2-5] and the resulting rejection of pollen occurs only if receptor and ligand are encoded by the same S haplotype [4, 6-8]. To identify residues within the SRK extracellular domain (eSRK) that are required for its ligand-selective activation, we assayed chimeric receptors and receptor variants containing substitutions at polymorphic sites in Arabidopsis thaliana[9, 10]. We show that only a small number of the approximately 100 polymorphic residues in eSRK are required for ligand-specific activation of self-incompatibility in vivo. These essential residues occur in two noncontiguous clusters located at equivalent positions in the two variants tested. They also correspond to sites showing elevated levels of substitutions in other SRKs, suggesting that these residues could define self-incompatibility specificity in most SRKs. The results demonstrate that the majority of eSRK residues that show signals of positive selection and previously surmised to function as specificity determinants are not essential for specificity in the SRK-SCR interaction.
Figures
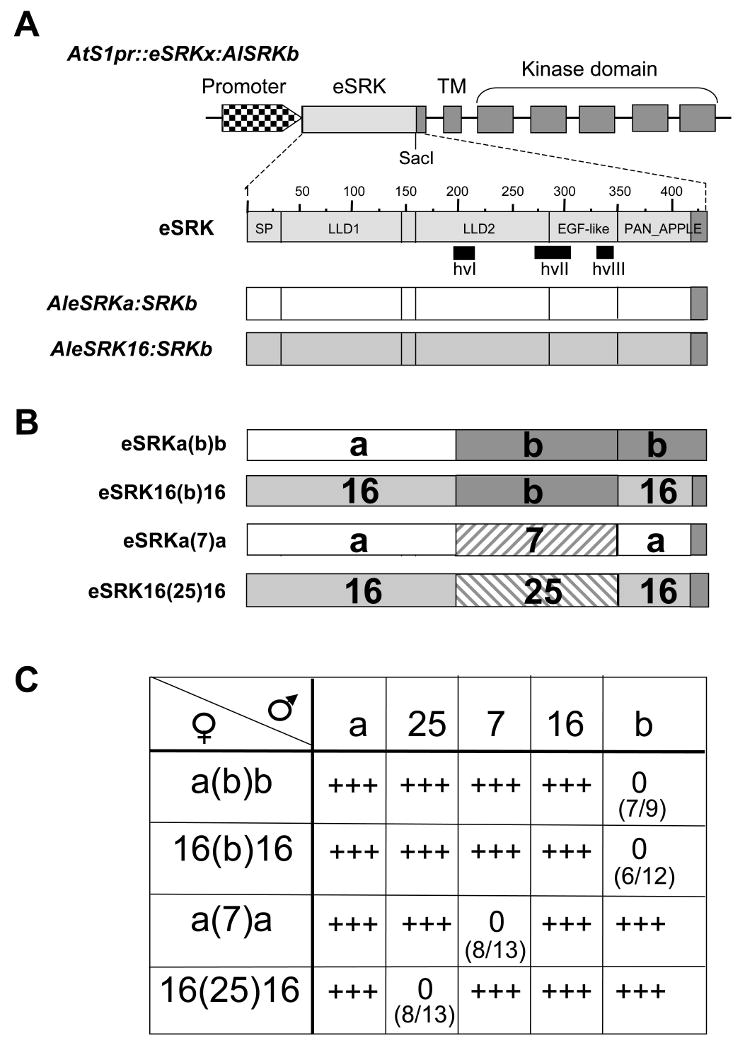
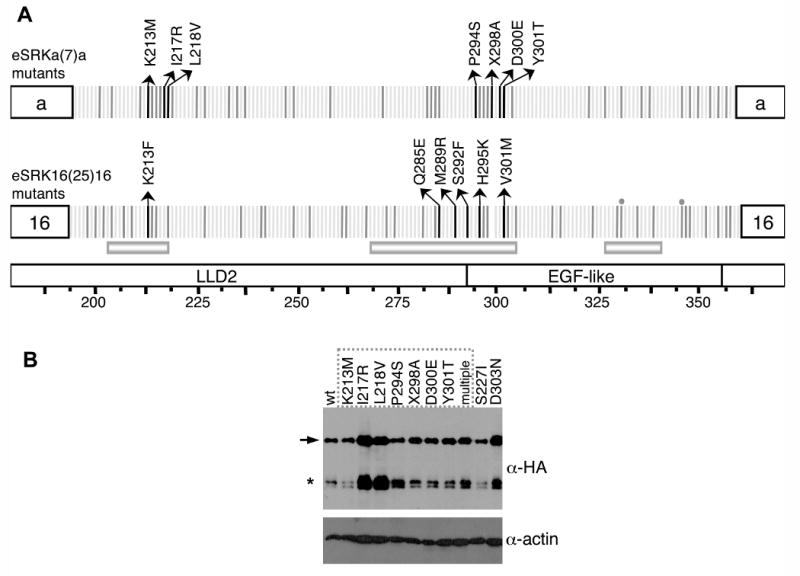
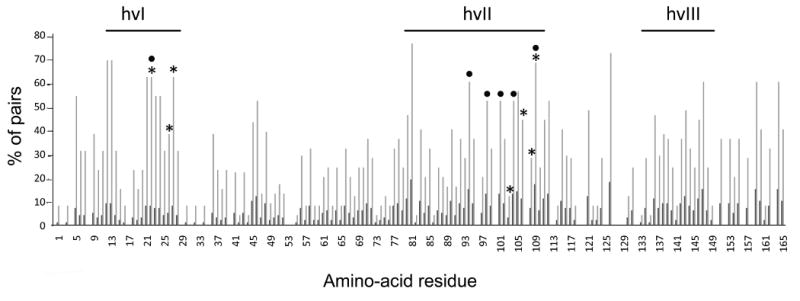
References
-
- Schopfer CR, Nasrallah ME, Nasrallah JB. The male determinant of self-incompatibility in Brassica. Science. 1999;286:1697–1700. - PubMed
-
- Kachroo A, Schopfer CR, Nasrallah ME, Nasrallah JB. Allele-specific receptor-ligand interactions in Brassica self-incompatibility. Science. 2001;293:1824–1826. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
