

PAT proteins, an ancient family of lipid droplet proteins that regulate cellular lipid stores
- PMID: 19375517
- PMCID: PMC2782626
- DOI: 10.1016/j.bbalip.2009.04.002
PAT proteins, an ancient family of lipid droplet proteins that regulate cellular lipid stores
Abstract
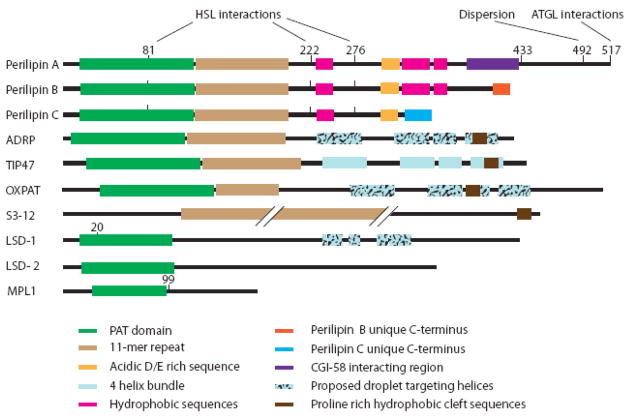
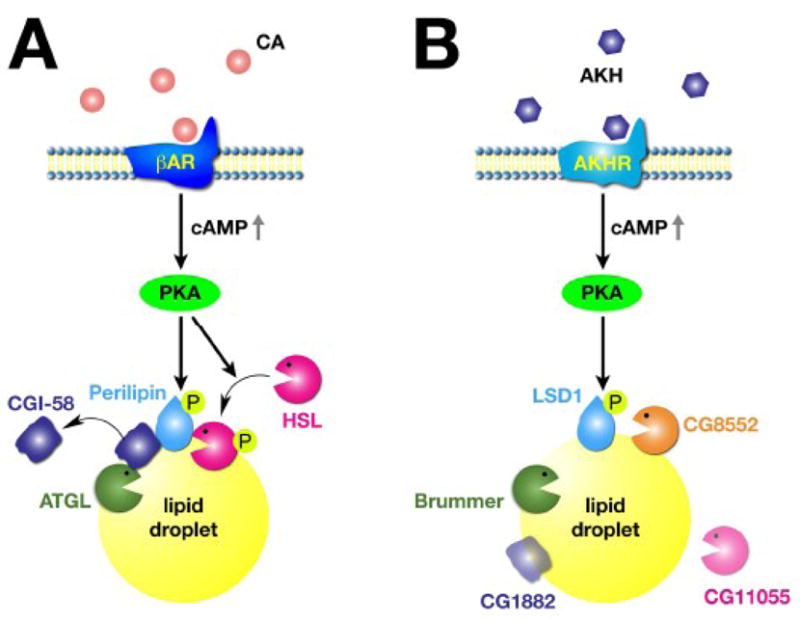
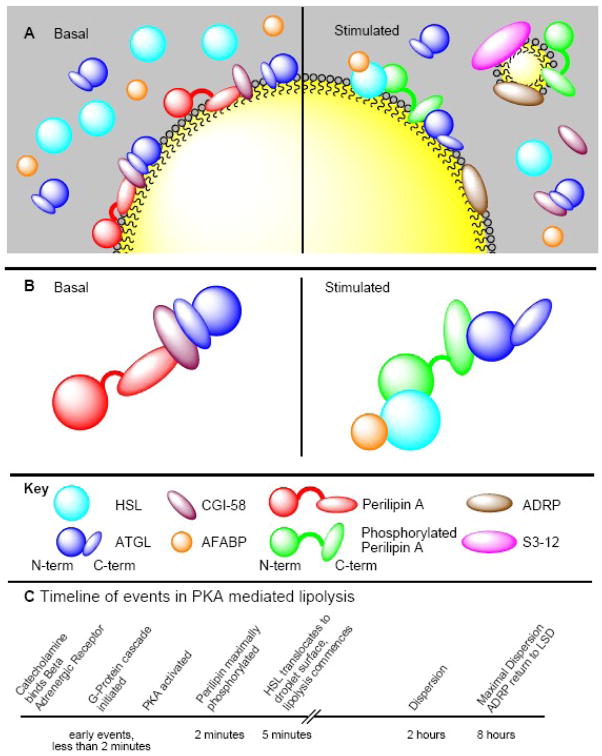
The PAT family of lipid droplet proteins includes 5 members in mammals: perilipin, adipose differentiation-related protein (ADRP), tail-interacting protein of 47 kDa (TIP47), S3-12, and OXPAT. Members of this family are also present in evolutionarily distant organisms, including insects, slime molds and fungi. All PAT proteins share sequence similarity and the ability to bind intracellular lipid droplets, either constitutively or in response to metabolic stimuli, such as increased lipid flux into or out of lipid droplets. Positioned at the lipid droplet surface, PAT proteins manage access of other proteins (lipases) to the lipid esters within the lipid droplet core and can interact with cellular machinery important for lipid droplet biogenesis. Genetic variations in the gene for the best-characterized of the mammalian PAT proteins, perilipin, have been associated with metabolic phenotypes, including type 2 diabetes mellitus and obesity. In this review, we discuss how the PAT proteins regulate cellular lipid metabolism both in mammals and in model organisms.
Figures
Similar articles
-
Adipocyte differentiation-related protein and OXPAT in rat and human skeletal muscle: involvement in lipid accumulation and type 2 diabetes mellitus.J Clin Endocrinol Metab. 2009 Oct;94(10):4077-85. doi: 10.1210/jc.2009-0352. Epub 2009 Jul 14. J Clin Endocrinol Metab. 2009. PMID: 19602560
-
Functional conservation for lipid storage droplet association among Perilipin, ADRP, and TIP47 (PAT)-related proteins in mammals, Drosophila, and Dictyostelium.J Biol Chem. 2002 Aug 30;277(35):32253-7. doi: 10.1074/jbc.M204410200. Epub 2002 Jun 20. J Biol Chem. 2002. PMID: 12077142
-
S3-12, Adipophilin, and TIP47 package lipid in adipocytes.J Biol Chem. 2005 May 13;280(19):19146-55. doi: 10.1074/jbc.M500978200. Epub 2005 Feb 24. J Biol Chem. 2005. PMID: 15731108
-
[Function of PAT family proteins in the lipid metabolism].Sheng Li Ke Xue Jin Zhan. 2006 Apr;37(2):103-7. Sheng Li Ke Xue Jin Zhan. 2006. PMID: 16850611 Review. Chinese.
-
Lipid droplet associated proteins: an emerging role in atherogenesis.Histol Histopathol. 2011 May;26(5):631-42. doi: 10.14670/HH-26.631. Histol Histopathol. 2011. PMID: 21432779 Review.
Cited by
-
ERα regulates lipid metabolism in bone through ATGL and perilipin.J Cell Biochem. 2013 Jun;114(6):1306-14. doi: 10.1002/jcb.24470. J Cell Biochem. 2013. PMID: 23296636 Free PMC article.
-
Hepatitis C virus comes for dinner: How the hepatitis C virus interferes with autophagy.World J Gastroenterol. 2015 Jul 28;21(28):8492-507. doi: 10.3748/wjg.v21.i28.8492. World J Gastroenterol. 2015. PMID: 26229393 Free PMC article. Review.
-
Polyunsaturated fatty acid relatively decreases cholesterol content in THP-1 macrophage-derived foam cell: partly correlates with expression profile of CIDE and PAT members.Lipids Health Dis. 2013 Jul 23;12:111. doi: 10.1186/1476-511X-12-111. Lipids Health Dis. 2013. PMID: 23879935 Free PMC article.
-
A multi-omic map of the lipid-producing yeast Rhodosporidium toruloides.Nat Commun. 2012;3:1112. doi: 10.1038/ncomms2112. Nat Commun. 2012. PMID: 23047670 Free PMC article.
-
Viperin catalyzes methionine oxidation to promote protein expression and function of helicases.Sci Adv. 2019 Aug 28;5(8):eaax1031. doi: 10.1126/sciadv.aax1031. eCollection 2019 Aug. Sci Adv. 2019. PMID: 31489375 Free PMC article.
References
-
- Murphy DJ. The biogenesis and functions of lipid bodies in animals, plants and microorganisms. Prog Lipid Res. 2001;40:325–438. - PubMed
-
- Bartz R, Zehmer JK, Zhu M, Chen Y, Serrero G, Zhao Y, Liu P. Dynamic activity of lipid droplets: protein phosphorylation and GTP-mediated protein translocation. J Proteome Res. 2007;6:3256–65. - PubMed
-
- Beller M, Riedel D, Jansch L, Dieterich G, Wehland J, Jackle H, Kuhnlein RP. Characterization of the Drosophila lipid droplet subproteome. Mol Cell Proteomics. 2006;5:1082–1094. - PubMed
-
- Brasaemle DL, Dolios G, Shapiro L, Wang R. Proteomic Analysis of Proteins Associated with Lipid Droplets of Basal and Lipolytically Stimulated 3T3-L1 Adipocytes. J Biol Chem. 2004;279:46835–42. - PubMed
-
- Liu P, Ying Y, Zhao Y, Mundy DI, Zhu M, Anderson RGW. Chinese Hamster Ovary K2 Cell Lipid Droplets Appear to Be Metabolic Organelles Involved in Membrane Traffic. J Biol Chem. 2004;279:3787–3792. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
