

Activation of the p75 neurotrophin receptor through conformational rearrangement of disulphide-linked receptor dimers
- PMID: 19376068
- PMCID: PMC2810632
- DOI: 10.1016/j.neuron.2009.02.020
Activation of the p75 neurotrophin receptor through conformational rearrangement of disulphide-linked receptor dimers
Abstract
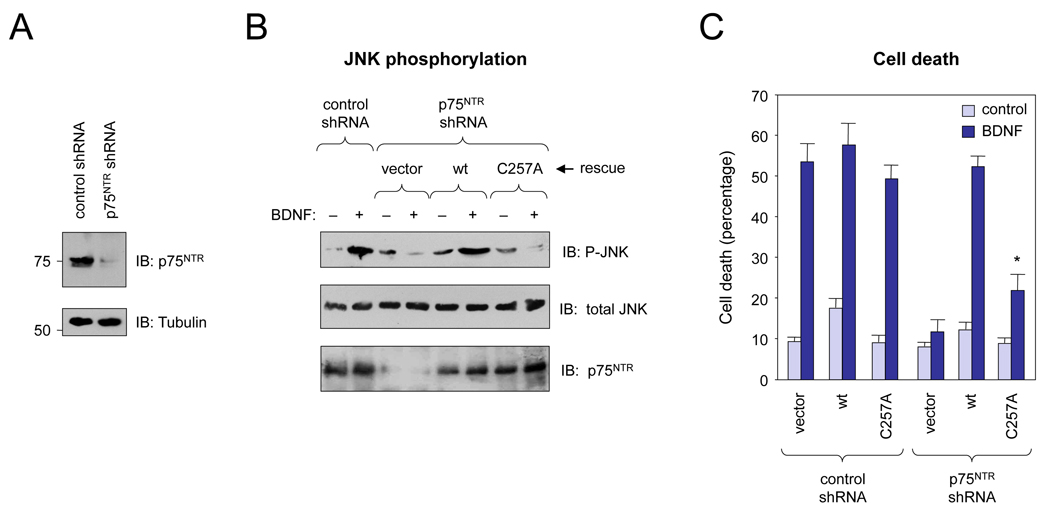
Ligand-mediated dimerization has emerged as a universal mechanism of growth factor receptor activation. Neurotrophins interact with dimers of the p75 neurotrophin receptor (p75(NTR)), but the mechanism of receptor activation has remained elusive. Here, we show that p75(NTR) forms disulphide-linked dimers independently of neurotrophin binding through the highly conserved Cys(257) in its transmembrane domain. Mutation of Cys(257) abolished neurotrophin-dependent receptor activity but did not affect downstream signaling by the p75(NTR)/NgR/Lingo-1 complex in response to MAG, indicating the existence of distinct, ligand-specific activation mechanisms for p75(NTR). FRET experiments revealed a close association of p75(NTR) intracellular domains that was transiently disrupted by conformational changes induced upon NGF binding. Although mutation of Cys(257) did not alter the oligomeric state of p75(NTR), the mutant receptor was no longer able to propagate conformational changes to the cytoplasmic domain upon ligand binding. We propose that neurotrophins activate p75(NTR) by a mechanism involving rearrangement of disulphide-linked receptor subunits.
Figures







Comment in
-
A p75(NTR) pivoting paradigm propels perspicacity.Neuron. 2009 Apr 16;62(1):3-5. doi: 10.1016/j.neuron.2009.04.005. Neuron. 2009. PMID: 19376060
References
-
- Ashkenazi A, Dixit VM. Apoptosis control by death and decoy receptors. Curr Opin Cell Biol. 1999;11:255–260. - PubMed
-
- Barker PA. p75NTR is positively promiscuous: novel partners and new insights. Neuron. 2004;42:529–533. - PubMed
-
- Bibel M, Barde YA. Neurotrophins: key regulators of cell fate and cell shape in the vertebrate nervous system. Genes Dev. 2000;14:2919–2937. - PubMed
-
- Bothwell M. Evolution of the neurotrophin signaling system in invertebrates. Brain Behav Evol. 2006;68:124–132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
