

Accurate sound localization in reverberant environments is mediated by robust encoding of spatial cues in the auditory midbrain
- PMID: 19376072
- PMCID: PMC2693331
- DOI: 10.1016/j.neuron.2009.02.018
Accurate sound localization in reverberant environments is mediated by robust encoding of spatial cues in the auditory midbrain
Abstract
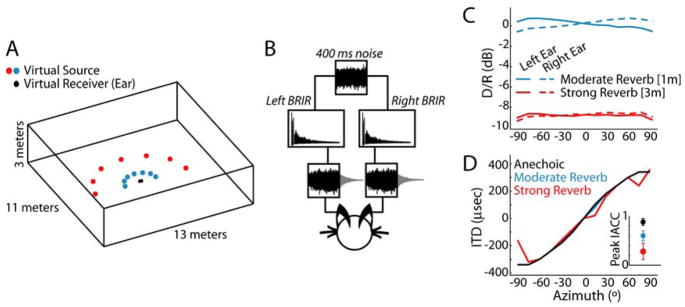
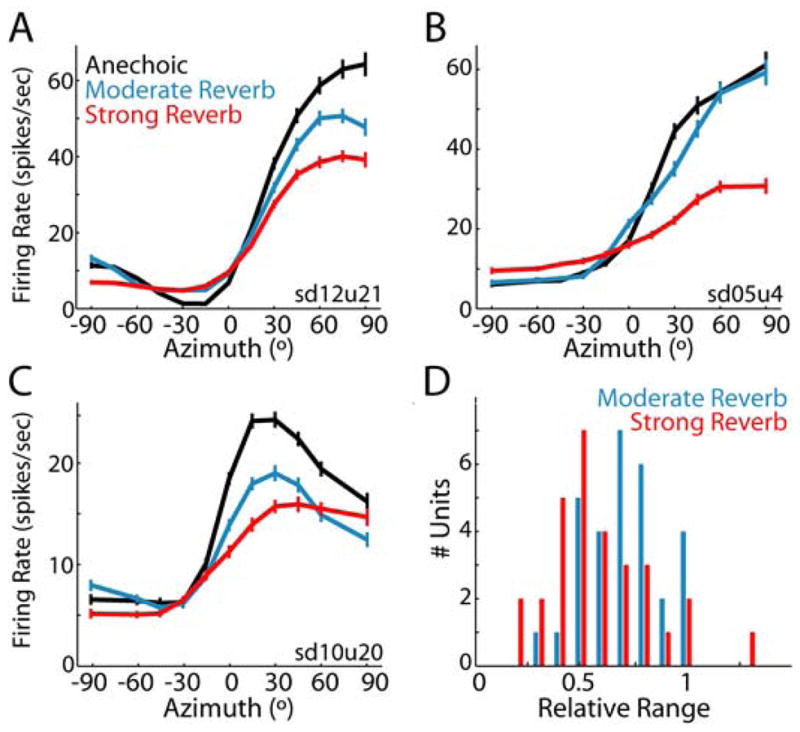
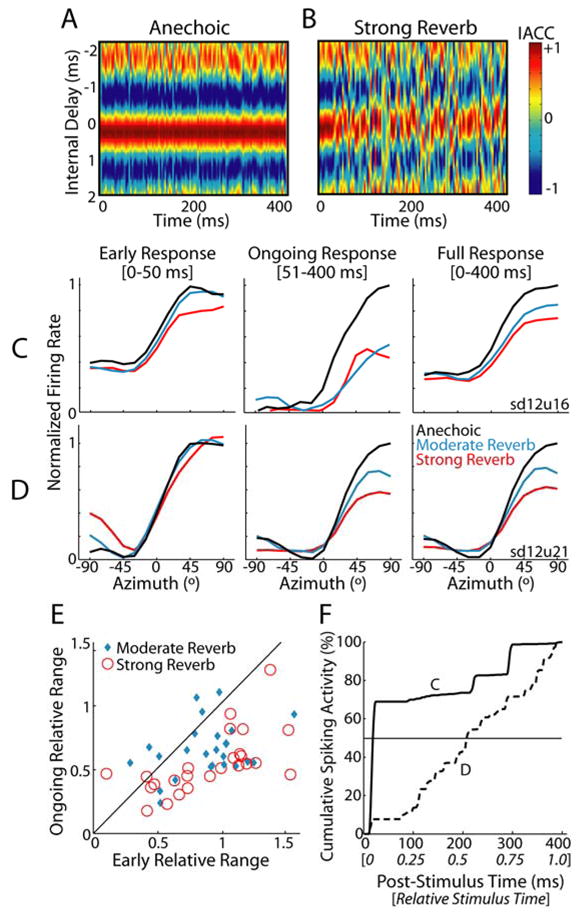
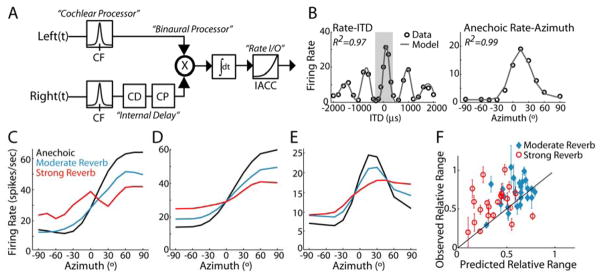
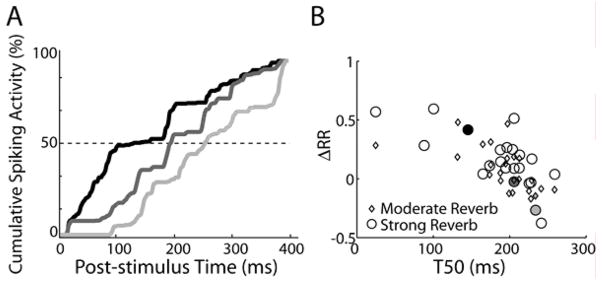
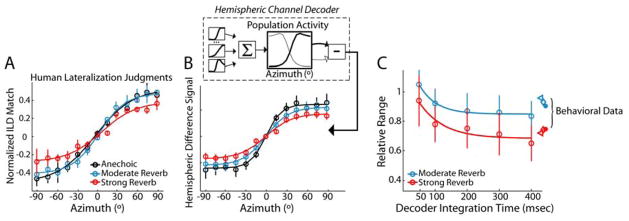
In reverberant environments, acoustic reflections interfere with the direct sound arriving at a listener's ears, distorting the spatial cues for sound localization. Yet, human listeners have little difficulty localizing sounds in most settings. Because reverberant energy builds up over time, the source location is represented relatively faithfully during the early portion of a sound, but this representation becomes increasingly degraded later in the stimulus. We show that the directional sensitivity of single neurons in the auditory midbrain of anesthetized cats follows a similar time course, although onset dominance in temporal response patterns results in more robust directional sensitivity than expected, suggesting a simple mechanism for improving directional sensitivity in reverberation. In parallel behavioral experiments, we demonstrate that human lateralization judgments are consistent with predictions from a population rate model decoding the observed midbrain responses, suggesting a subcortical origin for robust sound localization in reverberant environments.
Figures
Comment in
-
A novel view of hearing in reverberation.Neuron. 2009 Apr 16;62(1):6-7. doi: 10.1016/j.neuron.2009.04.004. Neuron. 2009. PMID: 19376061
References
-
- Adams JC. Ascending projections to the inferior colliculus. J Comp Neurol. 1979;183:519–538. - PubMed
-
- Aitkin LM, Gates GR, Phillips SC. Responses of Neurons in the Inferior Colliculus to Variations in Sound-Source Azimuth. Journal of Neurophysiology. 1984;52:1–17. - PubMed
-
- Albeck Y, Konishi M. Responses of neurons in the auditory pathway of the barn owl to partially correlated binaural signals. Journal of Neurophysiology. 1995;74:1689–1700. - PubMed
-
- Allen JB, Berkley DA. Image method for efficiently simulating small-room acoustics. J Acoust Soc Am. 1979;65:943–950.
-
- Beranek L. Concert Halls and Opera Houses. 2. New York: Springer-Verlag; 2004.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
