

The isomerase active site of cyclophilin A is critical for hepatitis C virus replication
- PMID: 19380579
- PMCID: PMC2719337
- DOI: 10.1074/jbc.M109.007625
The isomerase active site of cyclophilin A is critical for hepatitis C virus replication
Abstract
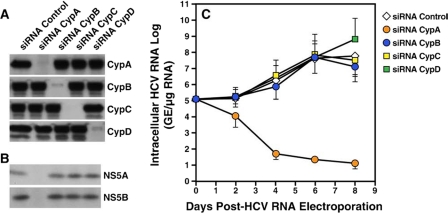
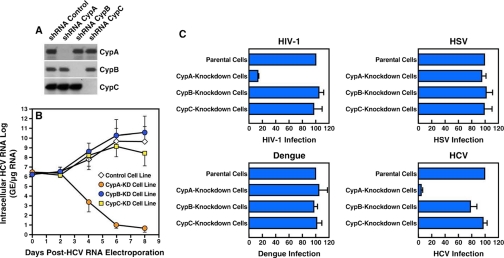
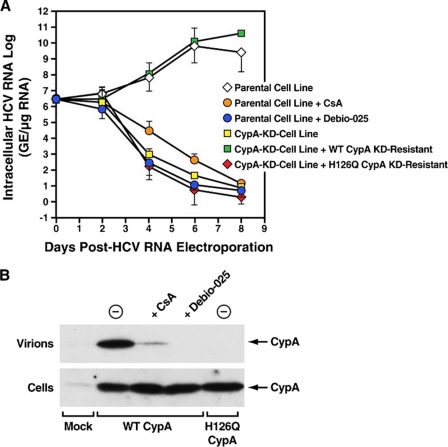
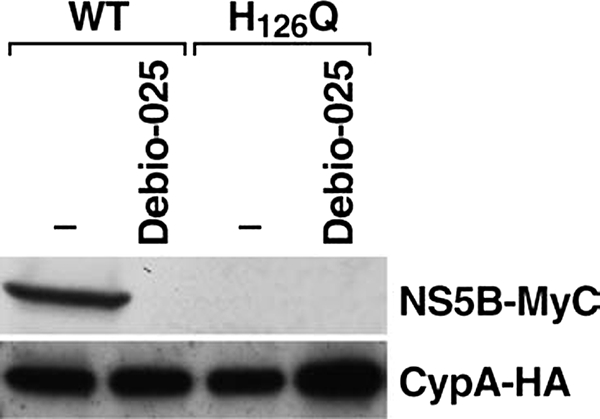
Cyclosporine A and nonimmunosuppressive cyclophilin (Cyp) inhibitors such as Debio 025, NIM811, and SCY-635 block hepatitis C virus (HCV) replication in vitro. This effect was recently confirmed in HCV-infected patients where Debio 025 treatment dramatically decreased HCV viral load, suggesting that Cyps inhibitors represent a novel class of anti-HCV agents. However, it remains unclear how these compounds control HCV replication. Recent studies suggest that Cyps are important for HCV replication. However, a profound disagreement currently exists as to the respective roles of Cyp members in HCV replication. In this study, we analyzed the respective contribution of Cyp members to HCV replication by specifically knocking down their expression by both transient and stable small RNA interference. Only the CypA knockdown drastically decreased HCV replication. The re-expression of an exogenous CypA escape protein, which contains escape mutations at the small RNA interference recognition site, restored HCV replication, demonstrating the specificity for the CypA requirement. We then mutated residues that reside in the hydrophobic pocket of CypA where proline-containing peptide substrates and cyclosporine A bind and that are vital for the enzymatic or the hydrophobic pocket binding activity of CypA. Remarkably, these CypA mutants fail to restore HCV replication, suggesting for the first time that HCV exploits either the isomerase or the chaperone activity of CypA to replicate in hepatocytes and that CypA is the principal mediator of the Cyp inhibitor anti-HCV activity. Moreover, we demonstrated that the HCV NS5B polymerase associates with CypA via its enzymatic pocket. The study of the roles of Cyps in HCV replication should lead to the identification of new targets for the development of alternate anti-HCV therapies.
Figures
References
-
- Pawlotsky J. M. ( 2006) Hepatology 43, S207– S220 - PubMed
-
- Hayashi N., Takehara T. ( 2006) J. Gastroenterol. 41, 17– 27 - PubMed
-
- Akiyama H., Yoshinaga H., Tanaka T., Hiruma K., Tanikawa S., Sakamaki H., Onozawa Y., Wakita T., Kohara M. ( 1997) Bone Marrow Transplant. 20, 993– 995 - PubMed
-
- Inoue K., Sekiyama K., Yamada M., Watanabe T., Yasuda H., Yoshiba M. ( 2003) J. Gastroenterol. 38, 567– 572 - PubMed
-
- Inoue K., Yoshiba M. ( 2005) Transplant. Proc. 37, 1233– 1234 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
