

H2O2-induced Ca2+ influx and its inhibition by N-(p-amylcinnamoyl) anthranilic acid in the beta-cells: involvement of TRPM2 channels
- PMID: 19382906
- PMCID: PMC4516483
- DOI: 10.1111/j.1582-4934.2009.00737.x
H2O2-induced Ca2+ influx and its inhibition by N-(p-amylcinnamoyl) anthranilic acid in the beta-cells: involvement of TRPM2 channels
Abstract
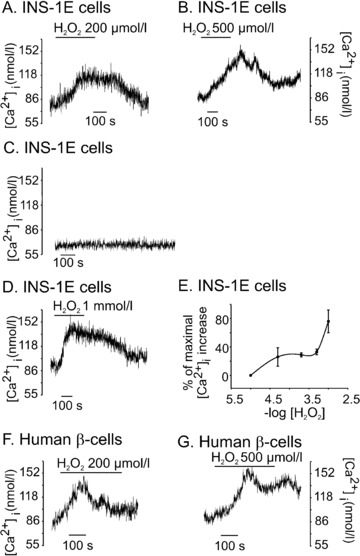
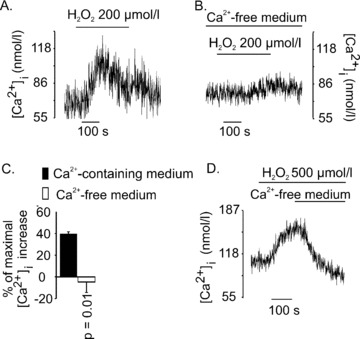
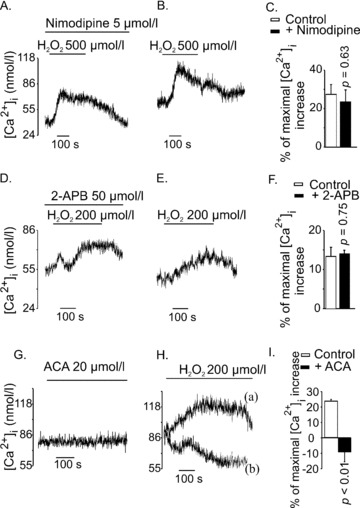
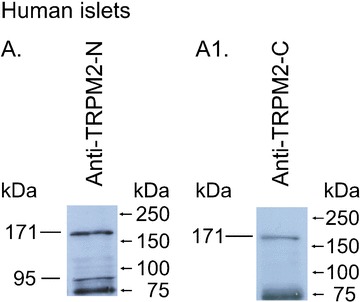
Type 2 melastatin-related transient receptor potential channel (TRPM2), a member of the melastatin-related TRP (transient receptor potential) subfamily is a Ca(2+)-permeable channel activated by hydrogen peroxide (H(2)O(2)). We have investigated the role of TRPM2 channels in mediating the H(2)O(2)-induced increase in the cytoplasmic free Ca(2+) concentration ([Ca(2+)](i)) in insulin-secreting cells. In fura-2 loaded INS-1E cells, a widely used model of beta-cells, and in human beta-cells, H(2)O(2) increased [Ca(2+)](i), in the presence of 3 mM glucose, by inducing Ca(2+) influx across the plasma membrane. H(2)O(2)-induced Ca(2+) influx was not blocked by nimodipine, a blocker of the L-type voltage-gated Ca(2+) channels nor by 2-aminoethoxydiphenyl borate, a blocker of several TRP channels and store-operated channels, but it was completely blocked by N-(p-amylcinnamoyl)anthranilic acid (ACA), a potent inhibitor of TRPM2. Adenosine diphosphate phosphate ribose, a specific activator of TRPM2 channel and H(2)O(2), induced inward cation currents that were blocked by ACA. Western blot using antibodies directed to the epitopes on the N-terminal and on the C-terminal parts of TRPM2 identified the full length TRPM2 (TRPM2-L), and the C-terminally truncated TRPM2 (TRPM2-S) in human islets. We conclude that functional TRPM2 channels mediate H(2)O(2)-induced Ca(2+) entry in beta-cells, a process potently inhibited by ACA.
Figures

References
-
- Perraud AL, Fleig A, Dunn CA, et al. ADP-ribose gating of the calcium-permeable LTRPC2 channel revealed by Nudix motif homology. Nature. 2001;411:595–9. - PubMed
-
- Maechler P, Jornot L, Wollheim CB. Hydrogen peroxide alters mitochondrial activation and insulin secretion in pancreatic beta cells. J Biol Chem. 1999;274:27905–13. - PubMed
-
- Pi J, Bai Y, Zhang Q, et al. Reactive oxygen species as a signal in glucose-stimulated insulin secretion. Diabetes. 2007;56:1783–91. - PubMed
-
- Qian F, Huang P, Ma L, et al. TRP genes: candidates for nonselective cation channels and store-operated channels in insulin-secreting cells. Diabetes. 2002;51:S183–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
