

Prestin's anion transport and voltage-sensing capabilities are independent
- PMID: 19383462
- PMCID: PMC2718310
- DOI: 10.1016/j.bpj.2008.12.3948
Prestin's anion transport and voltage-sensing capabilities are independent
Abstract
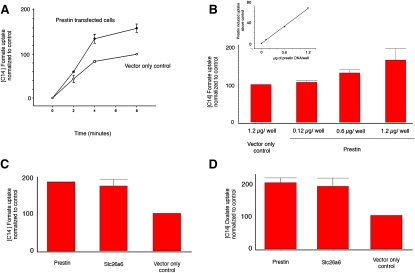
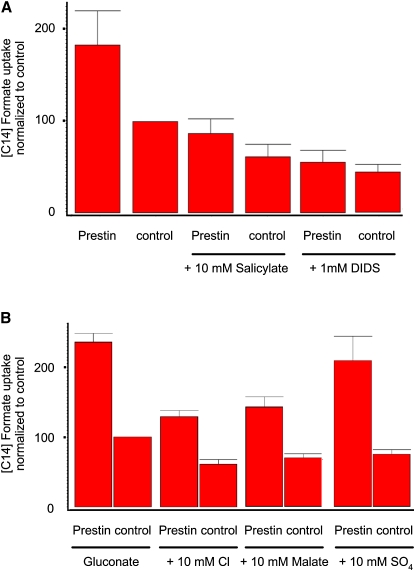
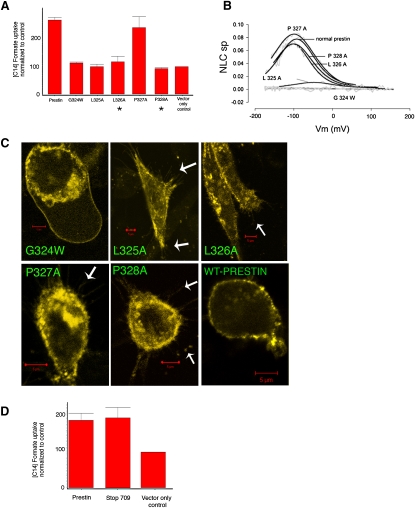
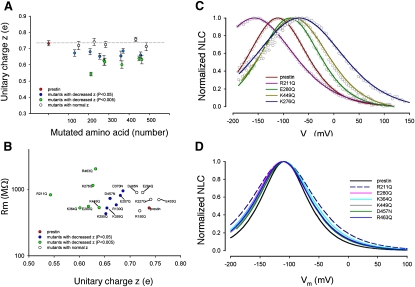
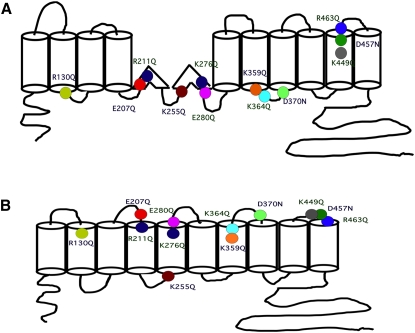
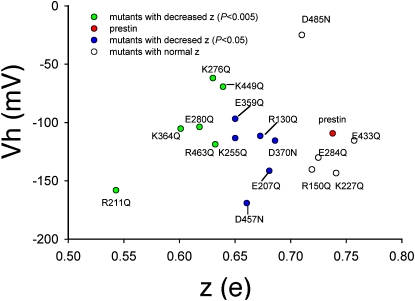
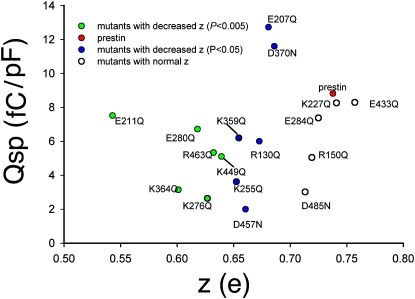
The integral membrane protein prestin, a member of the SLC26 anion transporter family, is responsible for the voltage-driven electromotility of mammalian outer hair cells. It was argued that the evolution of prestin's motor function required a loss of the protein's transport capabilities. Instead, it was proposed that prestin manages only an abortive hemicycle that results in the trapped anion acting as a voltage sensor, to generate the motor's signature gating charge movement or nonlinear capacitance. We demonstrate, using classical radioactive anion ([(14)C]formate and [(14)C]oxalate) uptake studies, that in contrast to previous observations, prestin is able to transport anions. The prestin-dependent uptake of both these anions was twofold that of cells transfected with vector alone, and comparable to SLC26a6, prestin's closest phylogenetic relative. Furthermore, we identify a potential chloride-binding site in which the mutations of two residues (P328A and L326A) preserve nonlinear capacitance, yet negate anion transport. Finally, we distinguish 12 charged residues out of 22, residing within prestin's transmembrane regions, that contribute to unitary charge movement, i.e., voltage sensing. These data redefine our mechanistic concept of prestin.
Figures
References
-
- Zheng J., Shen W., He D., Long K., Madison L. Prestin is the motor protein of cochlear outer hair cells. Nature. 2000;405:149–155. - PubMed
-
- Liberman M.C., Gao J., He D.Z., Wu X., Jia S. Prestin is required for electromotility of the outer hair cell and for the cochlear amplifier. Nature. 2002;419:300–304. - PubMed
-
- Reference deleted in proof.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
