

Muscle hypertrophy driven by myostatin blockade does not require stem/precursor-cell activity
- PMID: 19383783
- PMCID: PMC2671322
- DOI: 10.1073/pnas.0811129106
Muscle hypertrophy driven by myostatin blockade does not require stem/precursor-cell activity
Abstract
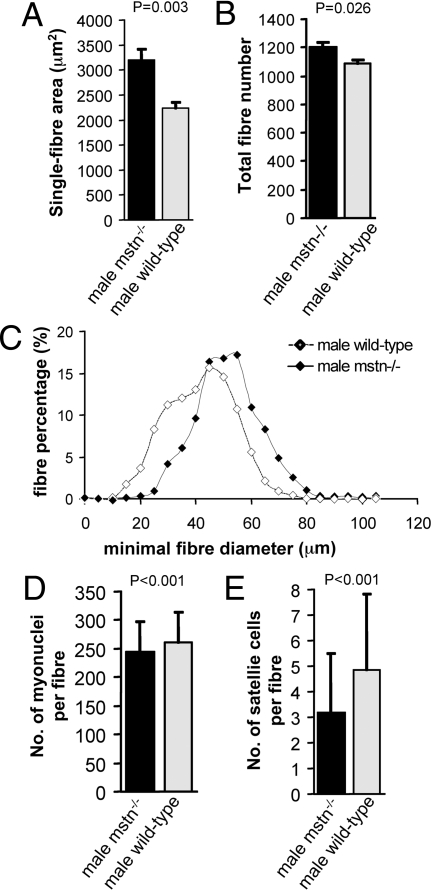
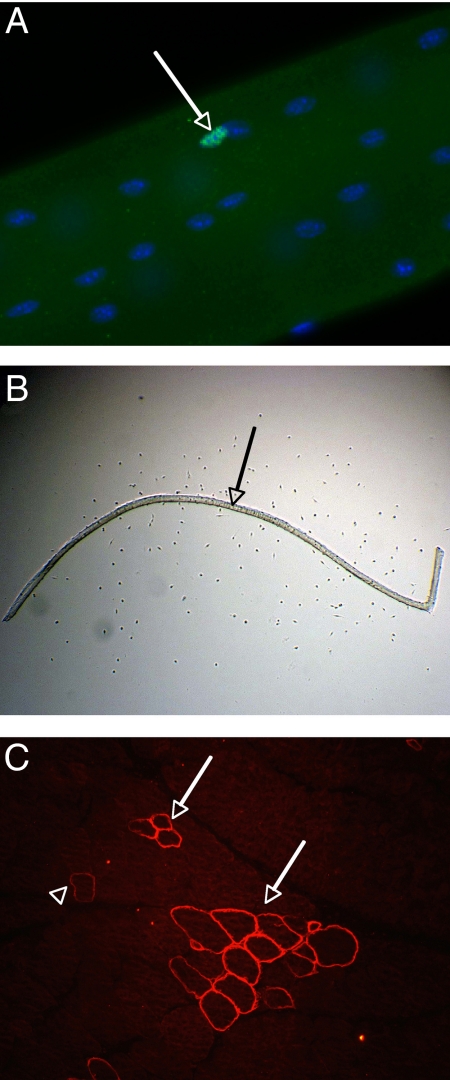
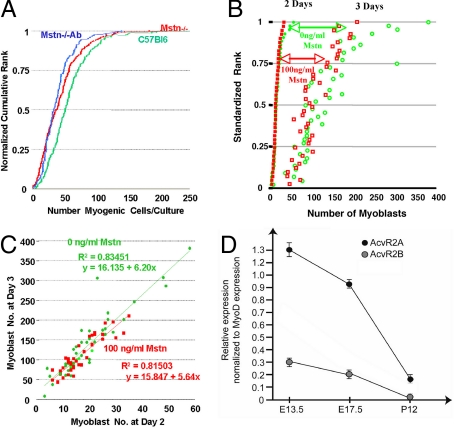
Myostatin, a member of the TGF-beta family, has been identified as a powerful inhibitor of muscle growth. Absence or blockade of myostatin induces massive skeletal muscle hypertrophy that is widely attributed to proliferation of the population of muscle fiber-associated satellite cells that have been identified as the principle source of new muscle tissue during growth and regeneration. Postnatal blockade of myostatin has been proposed as a basis for therapeutic strategies to combat muscle loss in genetic and acquired myopathies. But this approach, according to the accepted mechanism, would raise the threat of premature exhaustion of the pool of satellite cells and eventual failure of muscle regeneration. Here, we show that hypertrophy in the absence of myostatin involves little or no input from satellite cells. Hypertrophic fibers contain no more myonuclei or satellite cells and myostatin had no significant effect on satellite cell proliferation in vitro, while expression of myostatin receptors dropped to the limits of detectability in postnatal satellite cells. Moreover, hypertrophy of dystrophic muscle arising from myostatin blockade was achieved without any apparent enhancement of contribution of myonuclei from satellite cells. These findings contradict the accepted model of myostatin-based control of size of postnatal muscle and reorient fundamental investigations away from the mechanisms that control satellite cell proliferation and toward those that increase myonuclear domain, by modulating synthesis and turnover of structural muscle fiber proteins. It predicts too that any benefits of myostatin blockade in chronic myopathies are unlikely to impose any extra stress on the satellite cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- McPherron AC, Lawler AM, Lee S-J. Regulation of skeletal muscle mass in mice by a new TGF-β superfamily member. Nature. 1997;387:83–90. - PubMed
-
- Bogdanovich S, et al. Functional improvement of dystrophic muscle by myostatin blockade. Nature. 2002;420:418–421. - PubMed
-
- Wagner KR, McPherron AC, Winik N, Lee SJ. Loss of myostatin attenuates severity of muscular dystrophy in mdx mice. Ann Neurol. 2002;52:832–836. - PubMed
-
- McCroskery S, et al. Improved muscle healing through enhanced regeneration and reduced fibrosis in myostatin-null mice. J Cell Sci. 2005;118:3531–3541. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
