

Aberrant cleavage of TDP-43 enhances aggregation and cellular toxicity
- PMID: 19383787
- PMCID: PMC2671323
- DOI: 10.1073/pnas.0900688106
Aberrant cleavage of TDP-43 enhances aggregation and cellular toxicity
Abstract
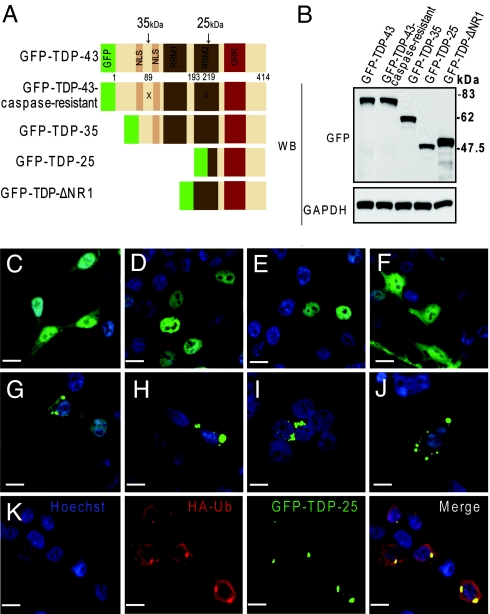
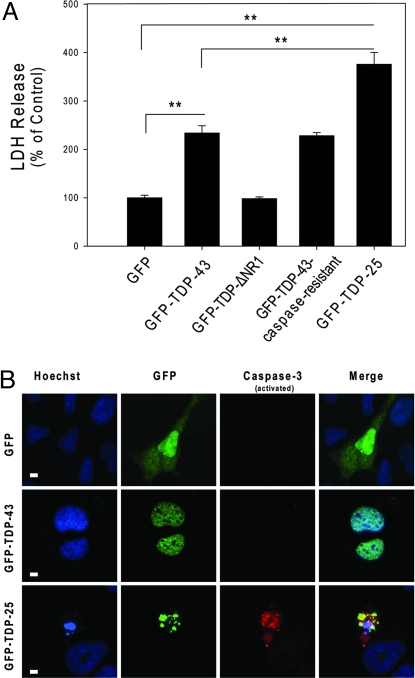
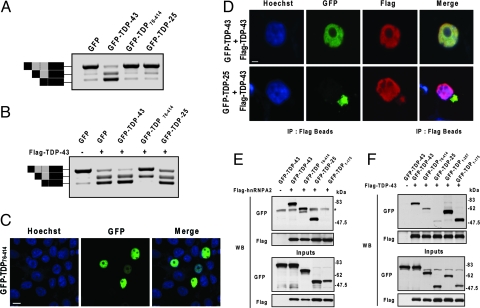
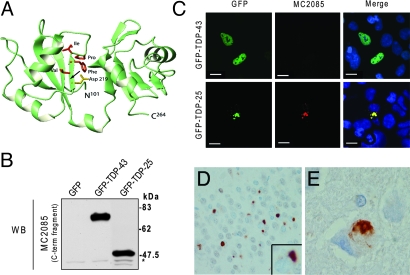
Inclusions of TAR DNA-binding protein-43 (TDP-43), a nuclear protein that regulates transcription and RNA splicing, are the defining histopathological feature of frontotemporal lobar degeneration with ubiquitin-positive inclusions (FTLD-Us) and sporadic and familial forms of amyotrophic lateral sclerosis (ALS). In ALS and FTLD-U, aggregated, ubiquitinated, and N-terminally truncated TDP-43 can be isolated from brain tissue rich in neuronal and glial cytoplasmic inclusions. The loss of TDP-43 function resulting from inappropriate cleavage, translocation from the nucleus, or its sequestration into inclusions could play important roles in neurodegeneration. However, it is not known whether TDP-43 fragments directly mediate toxicity and, more specifically, whether their abnormal aggregation is a cause or consequence of pathogenesis. We report that the ectopic expression of a approximately 25-kDa TDP-43 fragment corresponding to the C-terminal truncation product of caspase-cleaved TDP-43 leads to the formation of toxic, insoluble, and ubiquitin- and phospho-positive cytoplasmic inclusions within cells. The 25-kDa C-terminal fragment is more prone to phosphorylation at S409/S410 than full-length TDP-43, but phosphorylation at these sites is not required for inclusion formation or toxicity. Although this fragment shows no biological activity, its exogenous expression neither inhibits the function nor causes the sequestration of full-length nuclear TDP-43, suggesting that the 25-kDa fragment can induce cell death through a toxic gain-of-function. Finally, by generating a conformation-dependent antibody that detects C-terminal fragments, we show that this toxic cleavage product is specific for pathologic inclusions in human TDP-43 proteinopathies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Neumann M, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–133. - PubMed
-
- Arai T, et al. TDP-43 is a component of ubiquitin-positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Biochem Biophys Res Commun. 2006;351:602–611. - PubMed
-
- Dreyfuss G, Kim VN, Kataoka N. Messenger-RNA-binding proteins and the messages they carry. Nat Rev Mol Cell Biol. 2002;3:195–205. - PubMed
-
- Krecic AM, Swanson MS. hnRNP complexes: Composition, structure, and function. Curr Opin Cell Biol. 1999;11:363–371. - PubMed
-
- Buratti E, et al. TDP-43 binds heterogeneous nuclear ribonucleoprotein A/B through its C-terminal tail: An important region for the inhibition of cystic fibrosis transmembrane conductance regulator exon 9 splicing. J Biol Chem. 2005;280:37572–37584. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
