

ERalpha as ligand-independent activator of CDH-1 regulates determination and maintenance of epithelial morphology in breast cancer cells
- PMID: 19383788
- PMCID: PMC2671327
- DOI: 10.1073/pnas.0903033106
ERalpha as ligand-independent activator of CDH-1 regulates determination and maintenance of epithelial morphology in breast cancer cells
Abstract
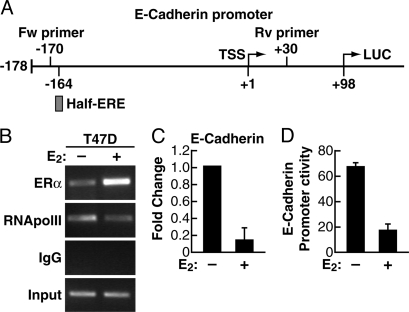
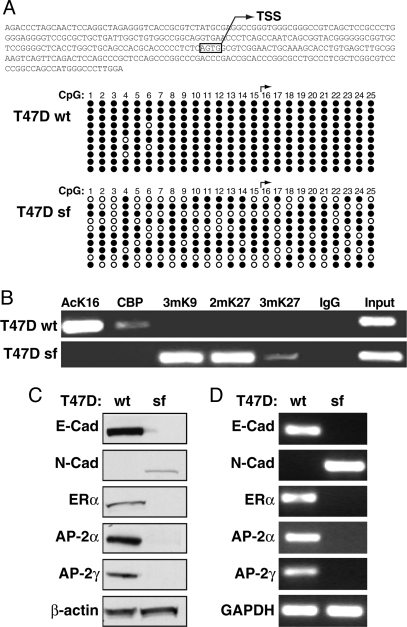
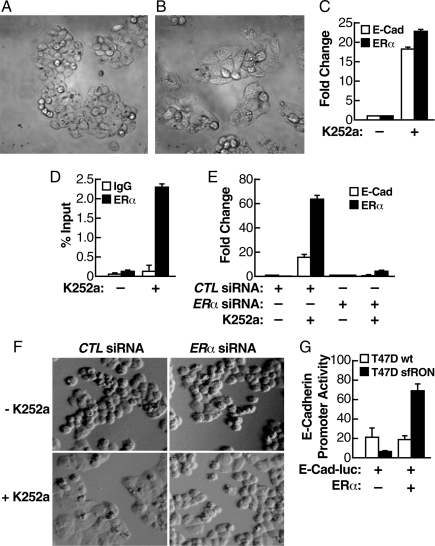
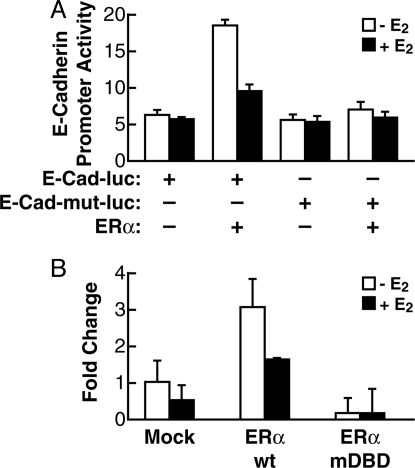
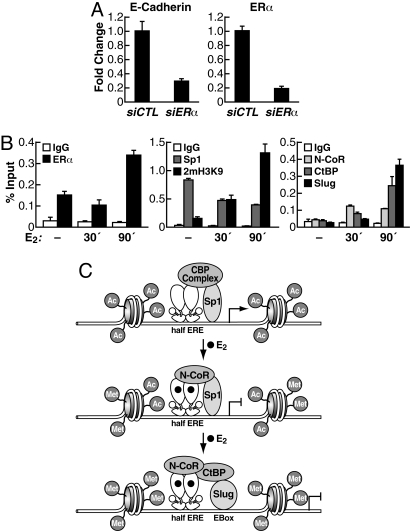
Estrogen receptor alpha (ERalpha) and E-cadherin are primary markers of luminal epithelial breast cancer cells with E-cadherin being a main caretaker of the epithelial phenotype. E-cadherin repression is needed for cancer cells to acquire motile and invasive properties, and it is known that in ER-positive breast cancer cells, estrogen down-regulate E-cadherin gene transcription. We report here that ERalpha is bound to the E-cadherin promoter in both the presence and the complete absence of estrogen, suggesting an unexpected role for unliganded ERalpha in E-cadherin transcription. Indeed, our data reveal that activation by unliganded ERalpha and repression by estrogen-activated ERalpha require direct binding to a half-estrogen response element within the E-cadherin promoter and exchange from associated coactivators to corepressors. Therefore, these results suggest a pivotal role for unliganded ERalpha in controlling a fundamental caretaker of the epithelial phenotype in breast cancer cells. Here, we show that ERalpha-positive breast cancer T47D cells transduced with the sfRON kinase undergo a full epithelial-mesenchymal conversion and lose E-cadherin and ERalpha expression. Our data show that, although the E-cadherin gene becomes hypermethylated and heterochromatic, kinase inhibitors can restore E-cadherin expression, together with an epithelial morphology in an ERalpha-dependent fashion. Similarly, transfection of ERalpha, in the absence of ligands, was sufficient to restore E-cadherin transcription in both sfRON-T47D and other ERalpha-, E-cadherin-negative cells. Therefore, our results suggest a novel role for the ERalpha that plays the dual role of ligand-independent activator and ligand-dependent repressor of E-cadherin in breast cancer cells.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
