

Mechanism of reduction in titers from lentivirus vectors carrying large inserts in the 3'LTR
- PMID: 19384292
- PMCID: PMC2835256
- DOI: 10.1038/mt.2009.89
Mechanism of reduction in titers from lentivirus vectors carrying large inserts in the 3'LTR
Abstract
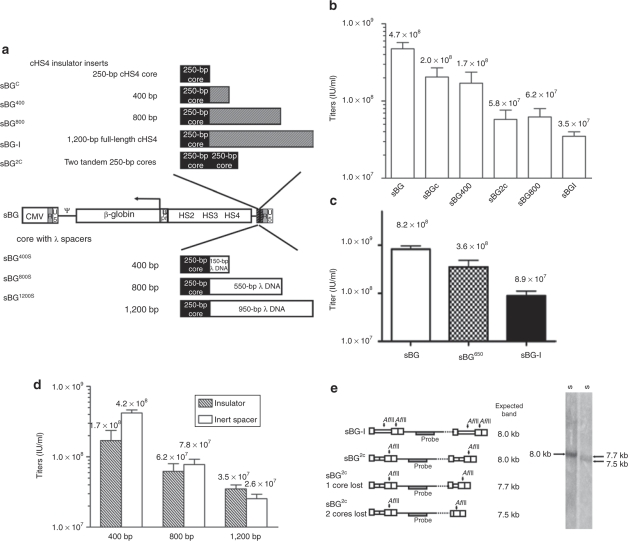
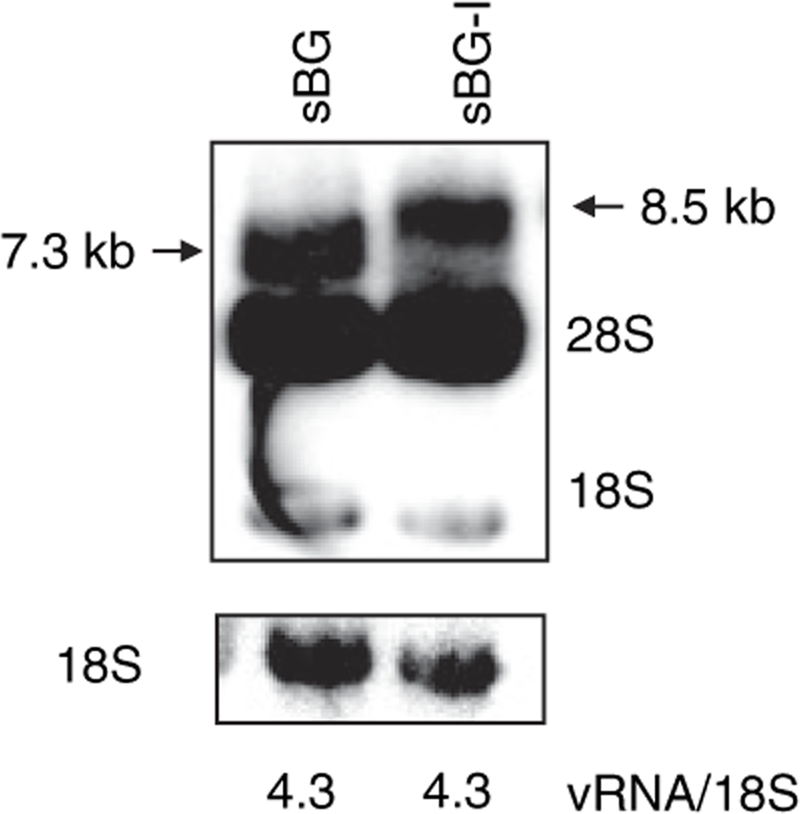
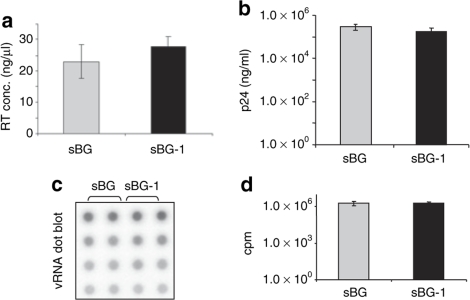
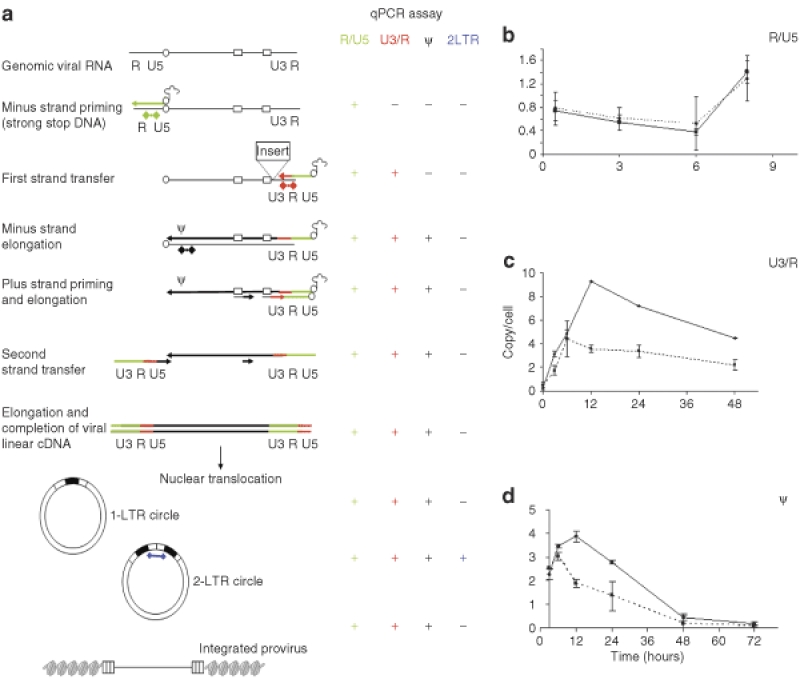
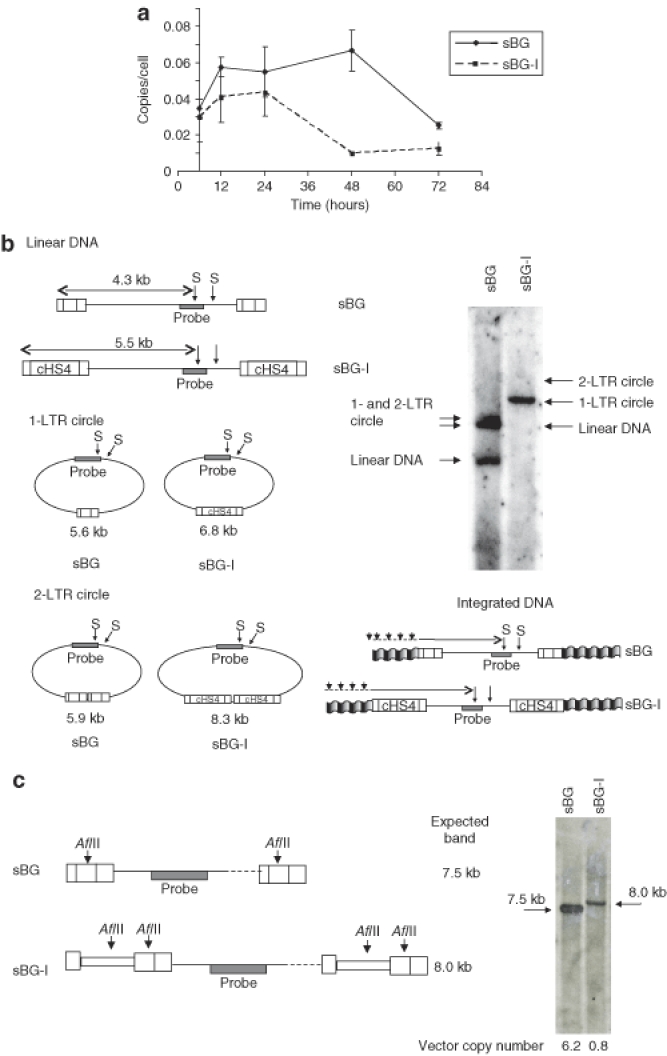
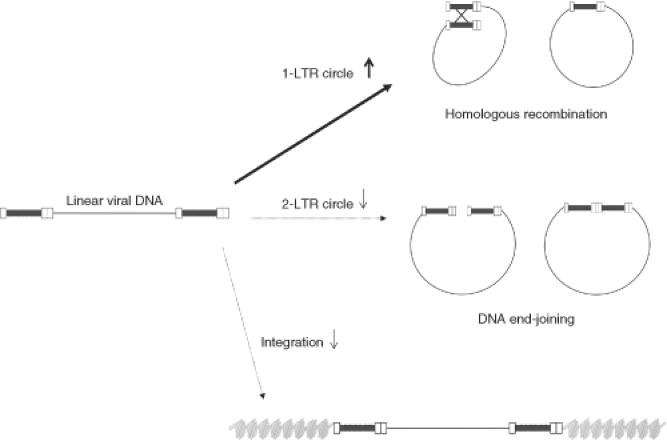
Self-inactivating (SIN) lentiviruses flanked by the 1.2-kb chicken hypersensitive site-4 (cHS4) insulator element provide consistent, improved expression of transgenes, but have significantly lower titers. The mechanism by which this occurs is unknown. Lengthening the lentiviral (LV) vector transgene cassette by an additional 1.2 kb by an internal cassette caused no further reduction in titers. However, when cHS4 sequences or inert DNA spacers of increasing size were placed in the 3'-long terminal repeat (LTR), infectious titers decreased proportional to the length of the insert. The stage of vector life cycle affected by vectors carrying the large cHS4 3'LTR insert was compared to a control vector: there was no increase in read-through transcription with insertion of the 1.2-kb cHS4 in the 3'LTR. Equal amount of full-length viral mRNA was produced in packaging cells and viral assembly/packaging was unaffected, resulting in comparable amounts of intact vector particles produced by either vectors. However, LV vectors carrying cHS4 in the 3'LTR were inefficiently processed following target-cell entry, with reduced reverse transcription and integration efficiency, and hence lower transduction titers. Therefore, vectors with large insertions in the 3'LTR are transcribed and packaged efficiently, but the LTR insert hinders viral-RNA (vRNA) processing and transduction of target cells. These studies have important implications in design of integrating vectors.
Figures
Similar articles
-
3' self-inactivating long terminal repeat inserts for the modulation of transgene expression from lentiviral vectors.Hum Gene Ther Methods. 2012 Apr;23(2):84-97. doi: 10.1089/hgtb.2011.154. Epub 2012 Mar 28. Hum Gene Ther Methods. 2012. PMID: 22456436
-
Advanced modular self-inactivating lentiviral expression vectors for multigene interventions in mammalian cells and in vivo transduction.Nucleic Acids Res. 2002 Nov 1;30(21):e113. doi: 10.1093/nar/gnf112. Nucleic Acids Res. 2002. PMID: 12409472 Free PMC article.
-
Large double copy vectors are functional but show a size-dependent decline in transduction efficiency.J Biotechnol. 2010 Oct 1;150(1):37-40. doi: 10.1016/j.jbiotec.2010.07.010. Epub 2010 Jul 16. J Biotechnol. 2010. PMID: 20638430
-
Successful correction of the human Cooley's anemia beta-thalassemia major phenotype using a lentiviral vector flanked by the chicken hypersensitive site 4 chromatin insulator.Ann N Y Acad Sci. 2005;1054:238-49. doi: 10.1196/annals.1345.030. Ann N Y Acad Sci. 2005. PMID: 16339671 Review.
-
Strategy and Application of Viral Potency Assays in Cell and Gene Therapy.Methods Mol Biol. 2025;2940:215-240. doi: 10.1007/978-1-0716-4615-1_20. Methods Mol Biol. 2025. PMID: 40515915 Review.
Cited by
-
CRISPR-Based Tools for Fighting Rare Diseases.Life (Basel). 2022 Nov 24;12(12):1968. doi: 10.3390/life12121968. Life (Basel). 2022. PMID: 36556333 Free PMC article. Review.
-
Potent microRNA suppression by RNA Pol II-transcribed 'Tough Decoy' inhibitors.RNA. 2013 Feb;19(2):280-93. doi: 10.1261/rna.034850.112. Epub 2012 Dec 18. RNA. 2013. PMID: 23249752 Free PMC article.
-
Coordinated β-globin expression and α2-globin reduction in a multiplex lentiviral gene therapy vector for β-thalassemia.Mol Ther. 2021 Sep 1;29(9):2841-2853. doi: 10.1016/j.ymthe.2021.04.037. Epub 2021 May 1. Mol Ther. 2021. PMID: 33940155 Free PMC article.
-
Modified gateway system for double shRNA expression and Cre/lox based gene expression.BMC Biotechnol. 2011 Mar 22;11:24. doi: 10.1186/1472-6750-11-24. BMC Biotechnol. 2011. PMID: 21418658 Free PMC article.
-
Improved Functionality of Integration-Deficient Lentiviral Vectors (IDLVs) by the Inclusion of IS2 Protein Docks.Pharmaceutics. 2021 Aug 6;13(8):1217. doi: 10.3390/pharmaceutics13081217. Pharmaceutics. 2021. PMID: 34452178 Free PMC article.
References
-
- Rivella S, May C, Chadburn A, Rivière I., and , Sadelain M. A novel murine model of Cooley anemia and its rescue by lentiviral-mediated human β-globin gene transfer. Blood. 2003;101:2932–2939. - PubMed
-
- Persons DA, Hargrove PW, Allay ER, Hanawa H., and , Nienhuis AW. The degree of phenotypic correction of murine β-thalassemia intermedia following lentiviral-mediated transfer of a human gamma-globin gene is influenced by chromosomal position effects and vector copy number. Blood. 2003;101:2175–2183. - PubMed
-
- Thrasher AJ, Gaspar HB, Baum C, Modlich U, Schambach A, Candotti F, et al. Gene therapy: X-SCID transgene leukaemogenicity. Nature. 2006;443:E5–6; discussion E6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
