

doi: 10.1002/dvdy.21951.
Increased Hox activity mimics the teratogenic effects of excess retinoic acid signaling
Affiliations
- PMID: 19384962
- PMCID: PMC2739864
- DOI: 10.1002/dvdy.21951
Item in Clipboard
Increased Hox activity mimics the teratogenic effects of excess retinoic acid signaling
Dev Dyn.
2009 May.
Abstract
Excess retinoic acid (RA) signaling can be teratogenic and result in cardiac birth defects, but the cellular and molecular origins of these defects are not well understood. Excessive RA signaling can completely eliminate heart formation in the zebrafish embryo. However, atrial and ventricular cells are differentially sensitive to more modest increases in RA signaling. Increased Hox activity, downstream of RA signaling, causes phenotypes similar to those resulting from excess RA. These results suggest that Hox activity mediates the differential effects of ectopic RA on atrial and ventricular cardiomyocytes and may underlie the teratogenic effects of RA on the heart.
Figures
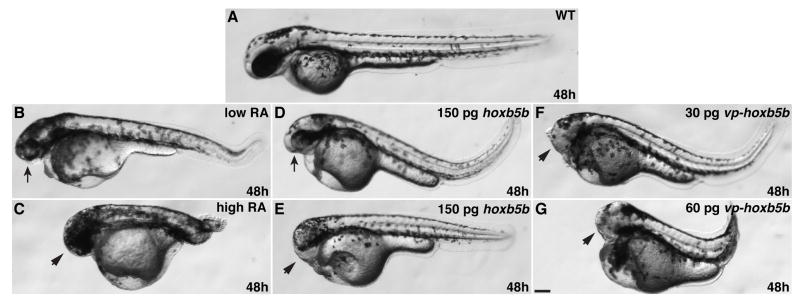
Increased RA signaling and Hox activity cause similar body axis phenotypes. (A) Wild-type embryo. (B) Low RA causes reduction of anterior head. (C) High RA causes loss of anterior head and tail truncation. (D and E) Injection of 150 pg of hoxb5b mRNA causes reduction or loss of anterior head. (F) 30 pg of vp-hoxb5b mRNA causes loss of anterior head. (G) 60 pg of vp-hoxb5b mRNA causes loss of anterior head and tail truncation. All images are at 48 hpf with anterior to the left. Arrows indicate reduced head and eyes. Arrowheads indicate loss of anterior head. Scale bar equals 100 μM.
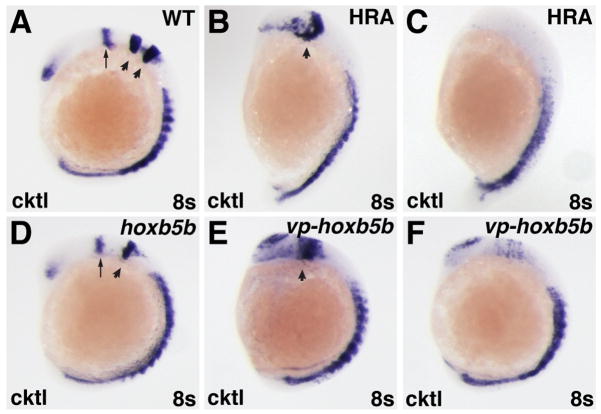
Increased RA signaling and Hox activity have similar effects on the anterior CNS. (A–F) In situ hybridizations at the 8-somite stage with a cocktail (cktl) of probes including pax2a (eye and MHB), krox-20/egr2b (rhombomeres 3 and 5), and myod (somites). (B,C,E,F) High RA treatment (HRA) or injection with 60 pg of vp-hoxb5b mRNA cause severe posteriorization phenotypes that vary between individual embryos. (B,E) High RA treatment and 60 pg vp-hoxb5b mRNA injection can truncate the anterior CNS, eliminate the MHB, and dramatically expand the posterior hindbrain. (C, F) High RA treatment and 60 pg vp-hoxb5b mRNA injection can also dramatically reduce or eliminate anterior CNS markers. (D) 150 pg hoxb5b mRNA injection causes loss of rhombomere 3, consistent with what has been shown previously for hoxb5b mRNA injection and for modest increases in RA signaling (Bruce et al., 2001; Hernandez et al., 2007). All images are lateral views, with dorsal to the right and anterior up. Arrows in A and D indicate MHB. Arrowheads in A indicate rhombomeres 3 and 5. Arrowheads in B, D, and E indicate rhombomere 5.
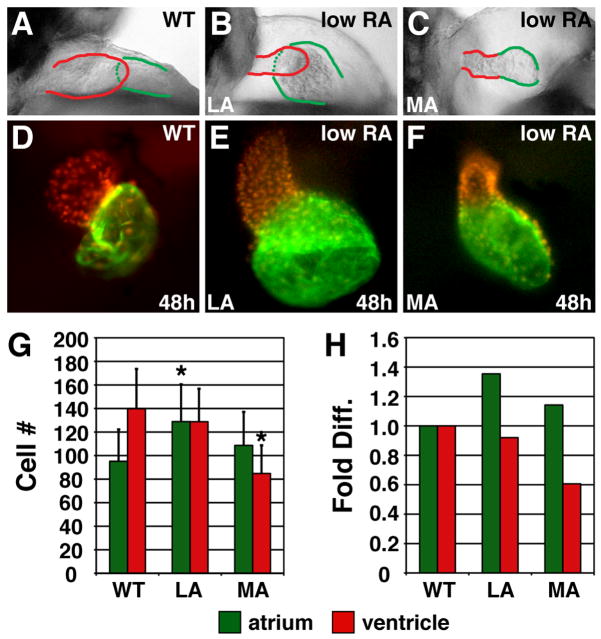
Excess RA signaling independently affects atrial and ventricular cell number. (A–C) Lateral views of hearts at 48 hpf; red and green outlines indicate ventricle and atrium, respectively. (D–F) Frontal views of Tg(cmlc2:DsRed2-nuc) (Mably et al., 2003) hearts, displaying nuclear DsRed in all cardiomyocytes, with Amhc immunofluorescence (green). (G) Mean (±SD) number of atrial and ventricular cardiomyocytes. WT, n=22; LA, n=45; MA, n=26. (H) Fold difference of the means in G. Asterisks indicate statistically significant differences from WT (p<0.005, Student’s t-test).
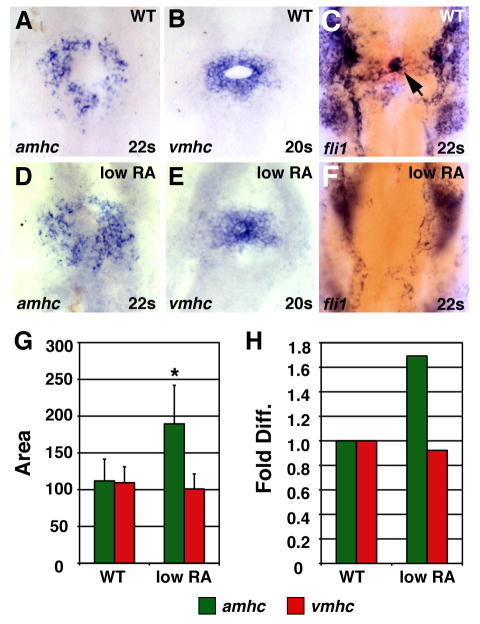
Low RA treatment can cause an increase in amhc expression. (A–F) In situ hybridizations at the 20 somite or 22 somite stage; dorsal views, anterior up. Arrow in C indicates endocardial precursors. (G) Mean (±SD) of areas of amhc and vmhc expression. WT amhc, n=43; WT vmhc, n=31; low RA amhc, n=18; low RA vmhc, n= 25. (H) Fold difference in means in G. Asterisk indicates statistically significant differences from WT (p<0.005, Student’s t-test).
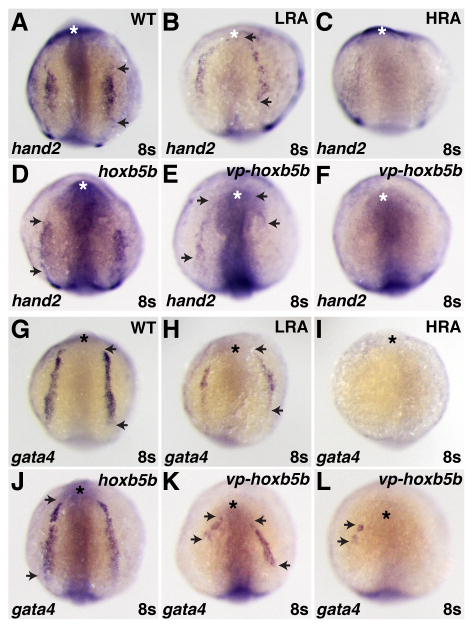
Increased RA signaling and Hox activity have similar effects on the ALPM. (A–L) In situ hybridizations of hand2 (A–F) and gata4 (G–L) in RA-treated, hoxb5b mRNA-injected, and vp-hoxb5b mRNA-injected embryos at the 8 somite stage. (D) 150 pg hoxb5b mRNA injection can cause a slight anterior shift in hand2 expression in the LPM. (B,E,H,K) Low RA (LRA) treatment or injection of 30 pg of vp-hoxb5b mRNA can shift hand2 anteriorly and reduce the length of gata4 expression. The uneven effects on either side of the embryo are likely due to mosaic distribution of the injected mRNA. (C,F,I,L) High RA (HRA) treatment or injection of 60 pg of vp-hoxb5b mRNA can eliminate hand2 and gata4 expression. All views are dorsal with anterior up. Asterisks indicate the anterior head. Arrowheads indicate the anterior and posterior borders of expression in the LPM.
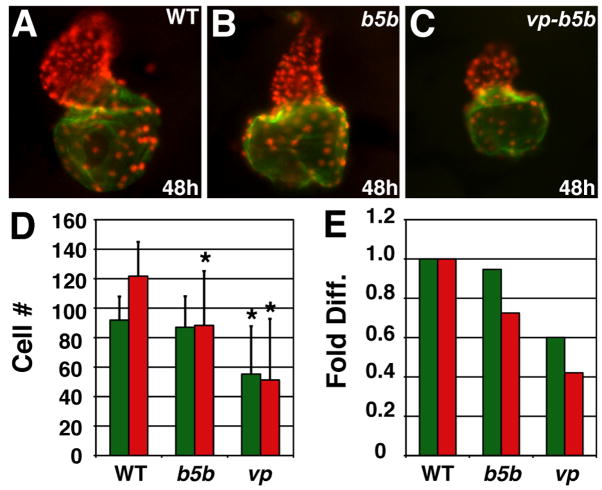
Increased Hox activity reduces atrial and ventricular cell number. (A–C) Hearts from WT, hoxb5b mRNA-injected (150 pg), and vp-hoxb5b mRNA-injected (60 pg) embryos. (D) Mean (±SD) number of atrial and ventricular cardiomyocytes. WT, n=25; hoxb5b mRNA, n=30; vp-hoxb5b mRNA, n=42. (E) Fold difference of means in D. Asterisks indicate statistically significant differences from WT (p<0.005, Student’s t-test). All views and graphs are as presented in Fig. 3.
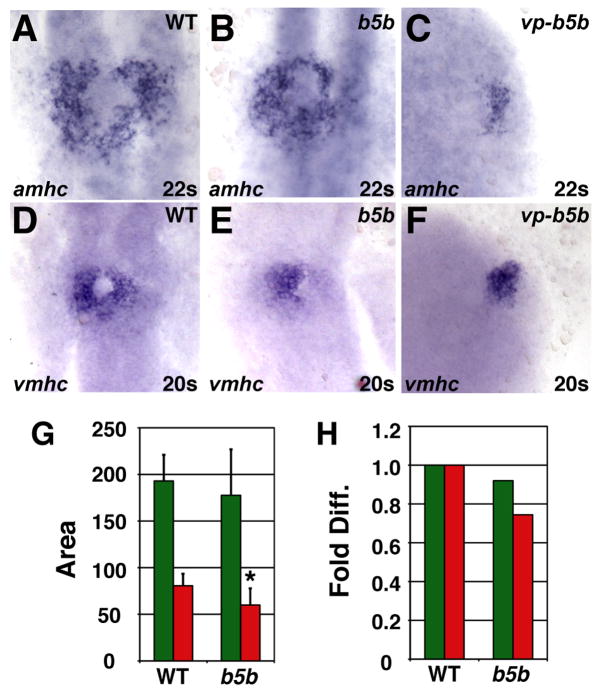
Increased Hox activity reduces amhc and vmhc expression. (A–F) Cardiomyocytes from WT, hoxb5b mRNA-injected (150 pg), and vp-hoxb5b mRNA-injected (60 pg) embryos. (G) Mean (±SD) of areas of amhc and vmhc expression. WT amhc, n=48; hoxb5b mRNA amhc, n=53; WT vmhc, n=59; hoxb5b mRNA vmhc, n=53. (H) Fold difference of means in G. Asterisk indicates statistically significant difference from WT (p<0.005, Student’s t-test). All views and graphs are as presented in Fig. 4.
References
-
- Berdougo E, Coleman H, Lee DH, Stainier DY, Yelon D. Mutation of weak atrium/atrial myosin heavy chain disrupts atrial function and influences ventricular morphogenesis in zebrafish. Development. 2003;130:6121–6129. - PubMed
-
- Bruce AE, Oates AC, Prince VE, Ho RK. Additional hox clusters in the zebrafish: divergent expression patterns belie equivalent activities of duplicate hoxB5 genes. Evol Dev. 2001;3:127–144. - PubMed
-
- Hochgreb T, Linhares VL, Menezes DC, Sampaio AC, Yan CY, Cardoso WV, Rosenthal N, Xavier-Neto J. A caudorostral wave of RALDH2 conveys anteroposterior information to the cardiac field. Development. 2003;130:5363–5374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
