

Apolipoprotein AI tertiary structures determine stability and phospholipid-binding activity of discoidal high-density lipoprotein particles of different sizes
- PMID: 19384992
- PMCID: PMC2771295
- DOI: 10.1002/pro.101
Apolipoprotein AI tertiary structures determine stability and phospholipid-binding activity of discoidal high-density lipoprotein particles of different sizes
Abstract
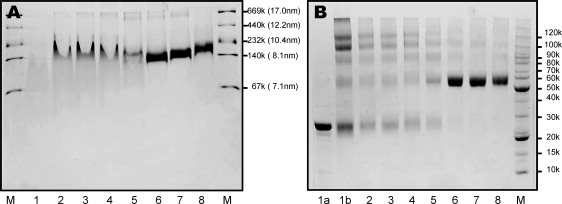
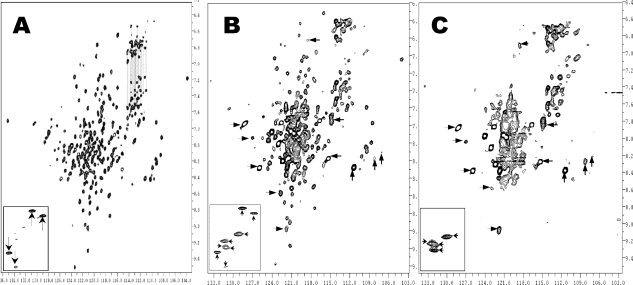
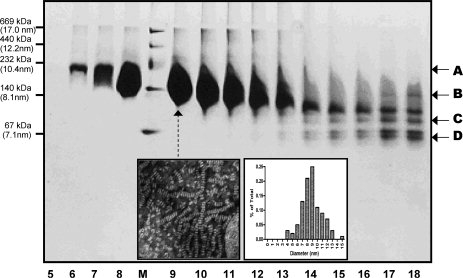
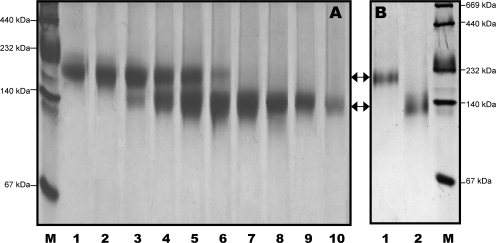
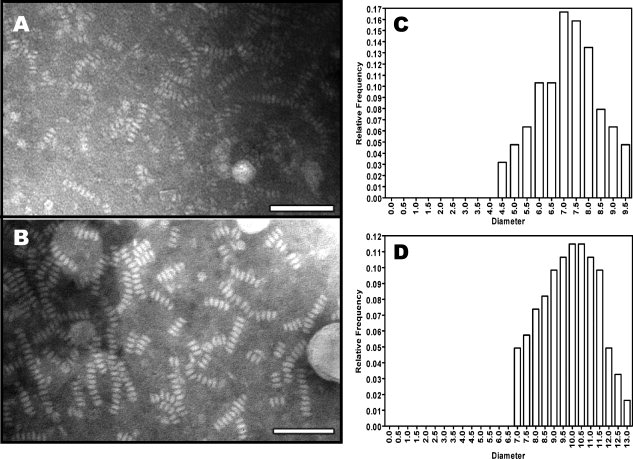
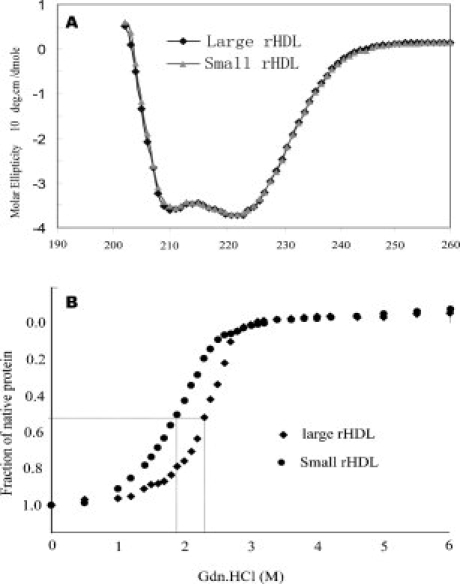
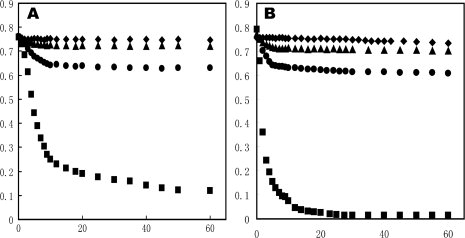
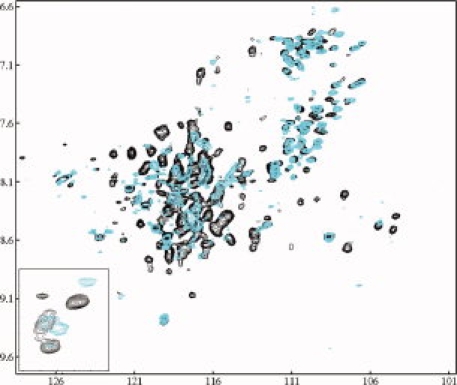
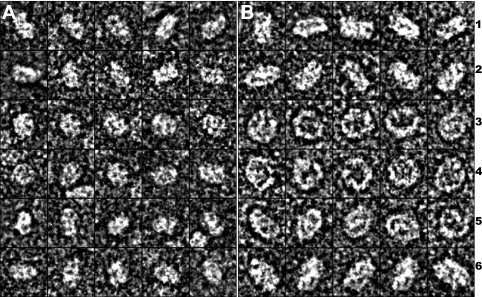
Human high-density lipoprotein (HDL) plays a key role in the reverse cholesterol transport pathway that delivers excess cholesterol back to the liver for clearance. In vivo, HDL particles vary in size, shape and biological function. The discoidal HDL is a 140-240 kDa, disk-shaped intermediate of mature HDL. During mature spherical HDL formation, discoidal HDLs play a key role in loading cholesterol ester onto the HDL particles by activating the enzyme, lecithin:cholesterol acyltransferase (LCAT). One of the major problems for high-resolution structural studies of discoidal HDL is the difficulty in obtaining pure and, foremost, homogenous sample. We demonstrate here that the commonly used cholate dialysis method for discoidal HDL preparation usually contains 5-10% lipid-poor apoAI that significantly interferes with the high-resolution structural analysis of discoidal HDL using biophysical methods. Using an ultracentrifugation method, we quickly removed lipid-poor apoAI. We also purified discoidal reconstituted HDL (rHDL) into two pure discoidal HDL species of different sizes that are amendable for high-resolution structural studies. A small rHDL has a diameter of 7.6 nm, and a large rHDL has a diameter of 9.8 nm. We show that these two different sizes of discoidal HDL particles display different stability and phospholipid-binding activity. Interestingly, these property/functional differences are independent from the apoAI alpha-helical secondary structure, but are determined by the tertiary structural difference of apoAI on different discoidal rHDL particles, as evidenced by two-dimensional NMR and negative stain electron microscopy data. Our result further provides the first high-resolution NMR data, demonstrating a promise of structural determination of discoidal HDL at atomic resolution using a combination of NMR and other biophysical techniques.
Figures
References
-
- Lewis GF, Rader DJ. New insights into the regulation of HDL metabolism and reverse cholesterol transport. Circ Res. 2005;96:1221–1232. - PubMed
-
- Davidson WS, Thompson TB. The structure of apolipoprotein a-I in high-density lipoproteins. J Biol Chem. 2007;282:22249–22253. - PubMed
-
- Asztalos BF. High-density lipoprotein metabolism and progression of atherosclerosis: new insights from the HDL Atherosclerosis Treatment Study. Curr Opin Cardiol. 2004;19:385–391. - PubMed
-
- Duriez P, Fruchart JC. High-density lipoprotein subclasses and apolipoprotein A-I. Clin Chim Acta. 1999;286:97–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
