

The SOCS box encodes a hierarchy of affinities for Cullin5: implications for ubiquitin ligase formation and cytokine signalling suppression
- PMID: 19385048
- PMCID: PMC2720833
- DOI: 10.1016/j.jmb.2009.01.024
The SOCS box encodes a hierarchy of affinities for Cullin5: implications for ubiquitin ligase formation and cytokine signalling suppression
Abstract
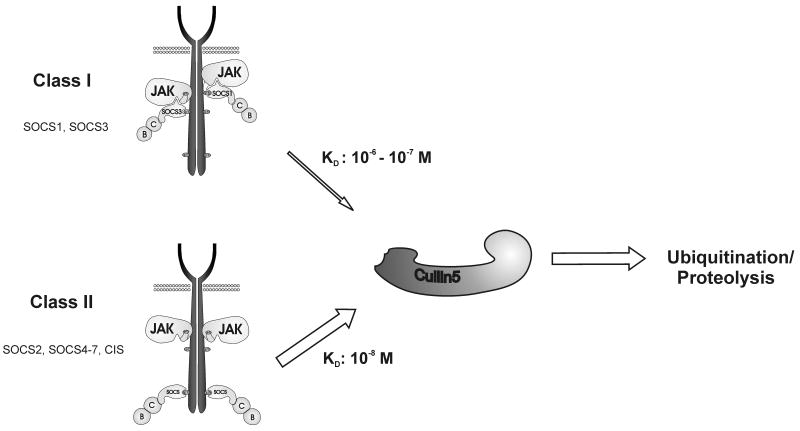
The SOCS (suppressors of cytokine signalling) family of proteins inhibits the cytokine-induced signalling cascade in part by promoting the ubiquitination of signalling intermediates that are then targeted for proteasomal degradation. This activity relies upon an interaction between the SOCS box domain, the adapter complex elonginBC and a member of the Cullin family, the scaffold protein of an E3 ubiquitin ligase. In this study, we dissected this interaction in vitro using purified components.We found that all eight SOCS proteins bound Cullin5 but required prior recruitment of elonginBC. Neither SOCS nor elonginBC bound Cullin5 when in isolation. Interestingly, the affinity of each SOCS-elonginBC complex for Cullin5 varied by 2 orders of magnitude across the SOCS family. Unexpectedly, the most potent suppressors of signalling, SOCS-1 and SOCS-3, bound most weakly to the E3 ligase scaffold, with affinities 100- and 10-fold lower, respectively, than the rest of the family. The remaining six SOCS proteins all bound Cullin5 with high affinity (K(d) of ~10 nM) due to a slower off-rate and hence a longer halflife of the complex. This difference in affinity may reflect a difference in mode of action as only SOCS-1 and SOCS-3 have been shown to suppress signalling using both SOCS box-dependent and SOCS box-independent mechanisms. This is not the case with the other six SOCS proteins, and our data imply the existence of two distinct subclasses of SOCS proteins with a high affinity for Cullin5, the E3 ligase scaffold, possibly reflecting complete dependence upon ubiquitination for suppression of cytokine signalling.
Figures





References
-
- Leonard WJ, O'Shea JJ. Jaks and STATs: biological implications. Annu Rev Immunol. 1998;16:293–322. - PubMed
-
- Starr R, Willson TA, Viney EM, Murray LJ, Rayner JR, Jenkins BJ, Gonda TJ, Alexander WS, Metcalf D, Nicola NA, Hilton DJ. A family of cytokine-inducible inhibitors of signalling. Nature. 1997;387:917–21. - PubMed
-
- Endo TA, Masuhara M, Yokouchi M, Suzuki R, Sakamoto H, Mitsui K, Matsumoto A, Tanimura S, Ohtsubo M, Misawa H, Miyazaki T, Leonor N, Taniguchi T, Fujita T, Kanakura Y, Komiya S, Yoshimura A. A new protein containing an SH2 domain that inhibits JAK kinases. Nature. 1997;387:921–4. - PubMed
-
- Naka T, Narazaki M, Hirata M, Matsumoto T, Minamoto S, Aono A, Nishimoto N, Kajita T, Taga T, Yoshizaki K, Akira S, Kishimoto T. Structure and function of a new STAT-induced STAT inhibitor. Nature. 1997;387:924–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
