

The importance of negative superhelicity in inducing the formation of G-quadruplex and i-motif structures in the c-Myc promoter: implications for drug targeting and control of gene expression
- PMID: 19385599
- PMCID: PMC2757002
- DOI: 10.1021/jm900055s
The importance of negative superhelicity in inducing the formation of G-quadruplex and i-motif structures in the c-Myc promoter: implications for drug targeting and control of gene expression
Abstract
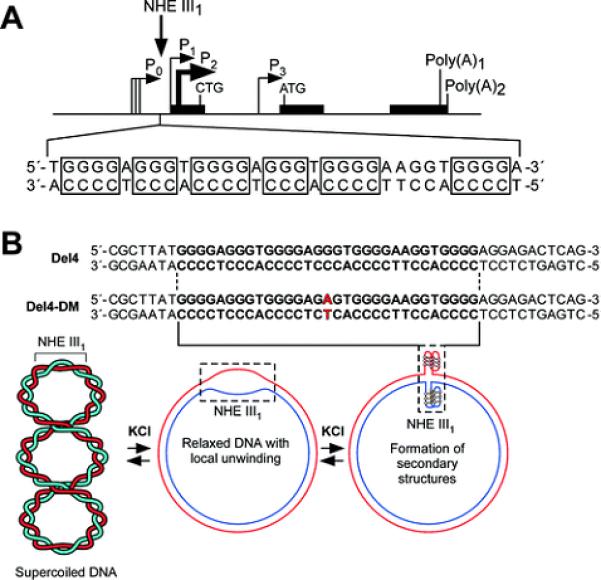
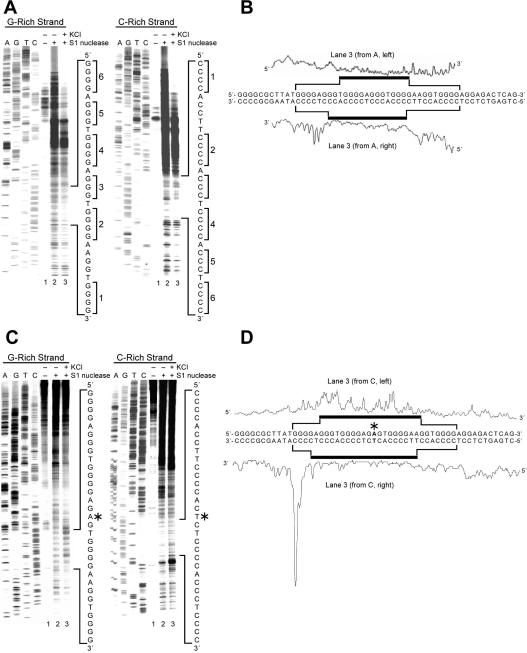
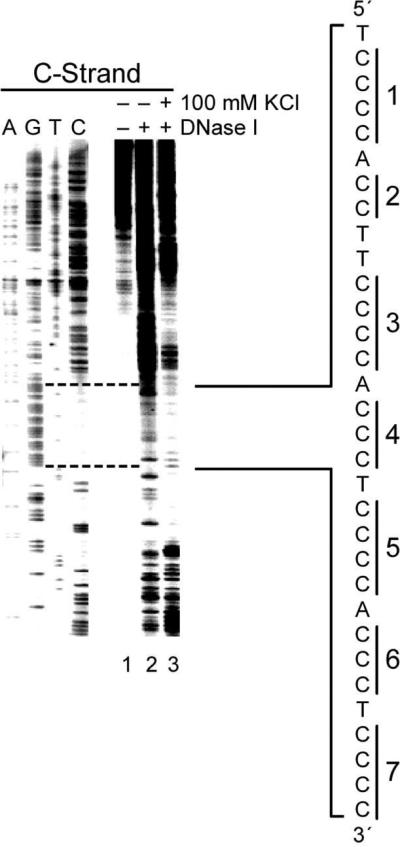
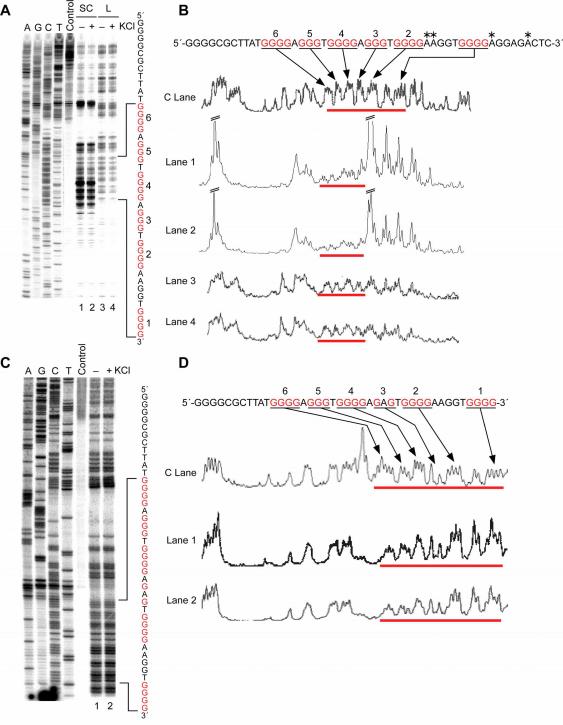
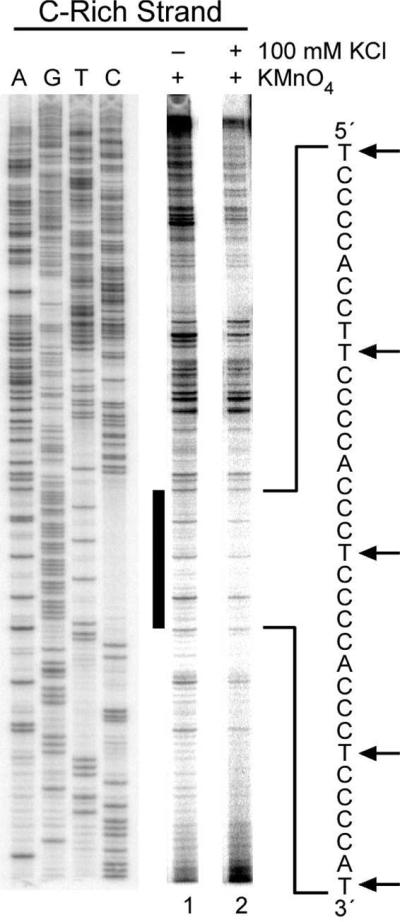
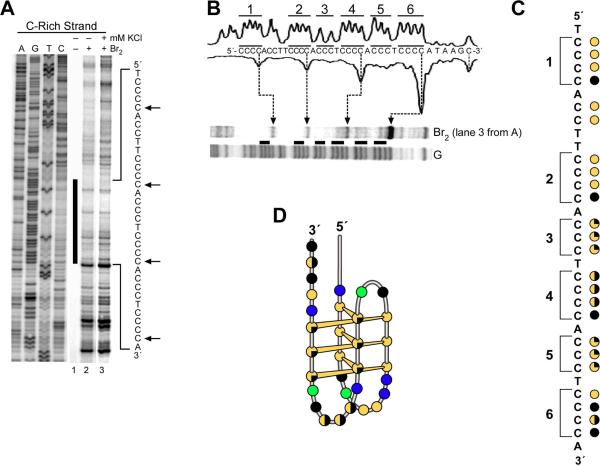
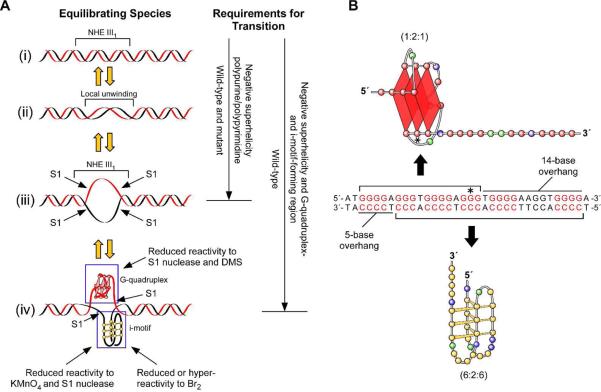
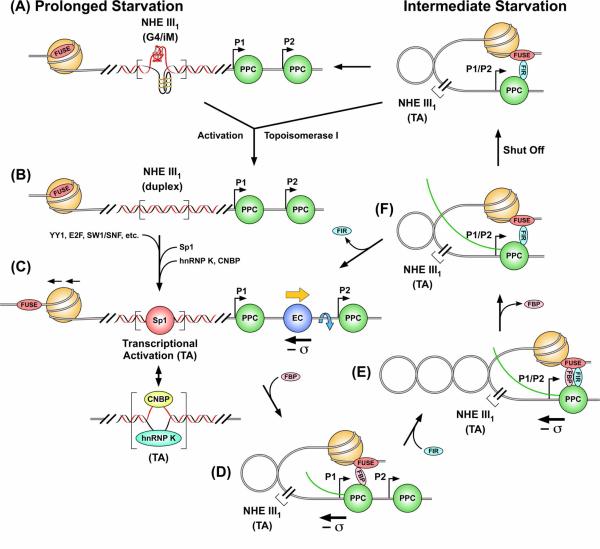
The importance of DNA supercoiling in transcriptional regulation has been known for many years, and more recently, transcription itself has been shown to be a source of this superhelicity. To mimic the effect of transcriptionally induced negative superhelicity, the G-quadruplex/i-motif-forming region in the c-Myc promoter was incorporated into a supercoiled plasmid. We show, using enzymatic and chemical footprinting, that negative superhelicity facilitates the formation of secondary DNA structures under physiological conditions. Significantly, these structures are not the same as those formed in single-stranded DNA templates. Together with the recently demonstrated role of transcriptionally induced superhelicity in maintaining a mechanosensor mechanism for controlling the firing rate of the c-Myc promoter, we provide a more complete picture of how c-Myc transcription is likely controlled. Last, these physiologically relevant G-quadruplex and i-motif structures, along with the mechanosensor mechanism for control of gene expression, are proposed as novel mechanisms for small molecule targeting of transcriptional control of c-Myc.
Figures
Similar articles
-
Facilitation of a structural transition in the polypurine/polypyrimidine tract within the proximal promoter region of the human VEGF gene by the presence of potassium and G-quadruplex-interactive agents.Nucleic Acids Res. 2005 Oct 20;33(18):6070-80. doi: 10.1093/nar/gki917. Print 2005. Nucleic Acids Res. 2005. PMID: 16239639 Free PMC article.
-
Multiple single-stranded cis elements are associated with activated chromatin of the human c-myc gene in vivo.Mol Cell Biol. 1996 Jun;16(6):2656-69. doi: 10.1128/MCB.16.6.2656. Mol Cell Biol. 1996. PMID: 8649373 Free PMC article.
-
NM23-H2 may play an indirect role in transcriptional activation of c-myc gene expression but does not cleave the nuclease hypersensitive element III(1).Mol Cancer Ther. 2009 May;8(5):1363-77. doi: 10.1158/1535-7163.MCT-08-1093. Epub 2009 May 12. Mol Cancer Ther. 2009. PMID: 19435876 Free PMC article.
-
The role of supercoiling in transcriptional control of MYC and its importance in molecular therapeutics.Nat Rev Cancer. 2009 Dec;9(12):849-61. doi: 10.1038/nrc2733. Epub 2009 Nov 12. Nat Rev Cancer. 2009. PMID: 19907434 Review.
-
Structure of the biologically relevant G-quadruplex in the c-MYC promoter.Nucleosides Nucleotides Nucleic Acids. 2006;25(8):951-68. doi: 10.1080/15257770600809913. Nucleosides Nucleotides Nucleic Acids. 2006. PMID: 16901825 Review.
Cited by
-
Opposite Effects of Potassium Ions on the Thermal Stability of i-Motif DNA in Different Buffer Systems.ACS Omega. 2021 Mar 24;6(13):8976-8985. doi: 10.1021/acsomega.0c06350. eCollection 2021 Apr 6. ACS Omega. 2021. PMID: 33842768 Free PMC article.
-
The roles of DNA methylation on pH dependent i-motif (iM) formation in rice.Nucleic Acids Res. 2024 Feb 9;52(3):1243-1257. doi: 10.1093/nar/gkad1245. Nucleic Acids Res. 2024. PMID: 38180820 Free PMC article.
-
Cleavage-defective Topoisomerase I mutants sharply increase G-quadruplex-associated genomic instability.Microb Cell. 2022 Jan 31;9(3):52-68. doi: 10.15698/mic2022.03.771. eCollection 2022 Mar 7. Microb Cell. 2022. PMID: 35291312 Free PMC article.
-
Remodeling the conformational dynamics of I-motif DNA by helicases in ATP-independent mode at acidic environment.iScience. 2021 Dec 6;25(1):103575. doi: 10.1016/j.isci.2021.103575. eCollection 2022 Jan 21. iScience. 2021. PMID: 34988409 Free PMC article.
-
Genomic Instability of G-Quadruplex Sequences in Escherichia coli: Roles of DinG, RecG, and RecQ Helicases.Genes (Basel). 2023 Aug 29;14(9):1720. doi: 10.3390/genes14091720. Genes (Basel). 2023. PMID: 37761860 Free PMC article.
References
-
- Marcu KB. Regulation of expression of the c-myc proto-oncogene. Bioessays. 1987;6:28–32. - PubMed
-
- Pelengaris S, Khan M, Evan GI. Suppression of Myc-induced apoptosis in β cells exposes multiple oncogenic properties of Myc and triggers carcinogenic progression. Cell. 2002;109:321–334. - PubMed
-
- Pelengaris S, Khan M. The c-Myc oncoprotein as a treatment target in cancer and other disorders of cell growth. Expert Opin. Ther. Targets. 2003;7:623–642. - PubMed
-
- Spencer CA, Groudine M. Control of c-myc regulation in normal and neoplastic cells. Adv. Cancer Res. 1991;56:1–48. - PubMed
-
- Marcu KB, Patel AJ, Yang Y. Differential regulation of the c-Myc P1 and P2 promoters in the absence of functional tumor suppressors: implications for mechanisms of deregulated Myc transcription. Curr. Top. Microbiol. Immunol. 1997;224:47–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
