

Immunomodulatory parasites and toll-like receptor-mediated tumour necrosis factor alpha responsiveness in wild mammals
- PMID: 19386086
- PMCID: PMC2685781
- DOI: 10.1186/1741-7007-7-16
Immunomodulatory parasites and toll-like receptor-mediated tumour necrosis factor alpha responsiveness in wild mammals
Abstract
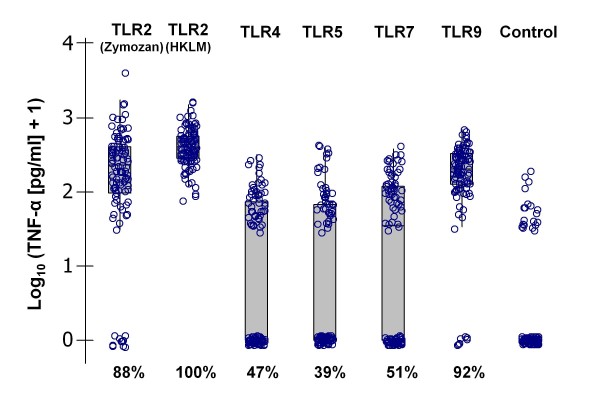
Background: Immunological analyses of wild populations can increase our understanding of how vertebrate immune systems respond to 'natural' levels of exposure to diverse infections. A major recent advance in immunology has been the recognition of the central role of phylogenetically conserved toll-like receptors in triggering innate immunity and the subsequent recruitment of adaptive response programmes. We studied the cross-sectional associations between individual levels of systemic toll-like receptor-mediated tumour necrosis factor alpha responsiveness and macro- and microparasite infections in a natural wood mouse (Apodemus sylvaticus) population.
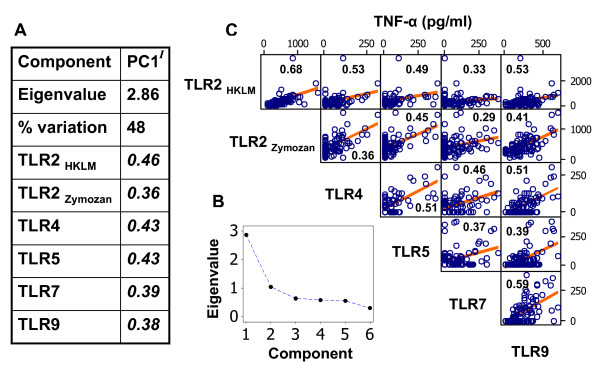
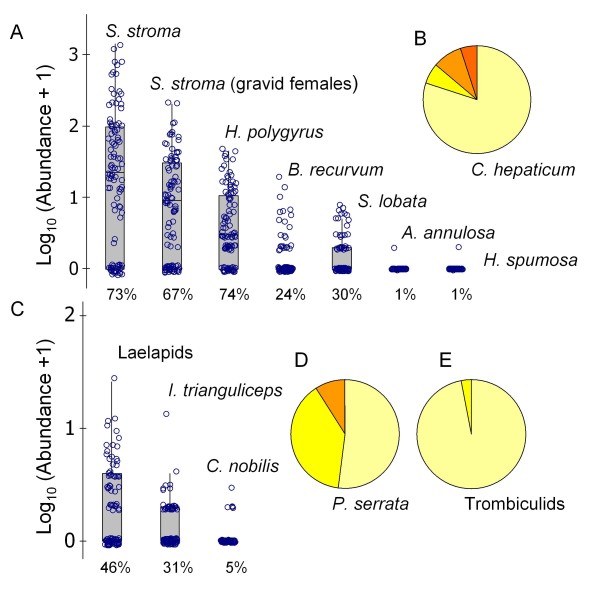
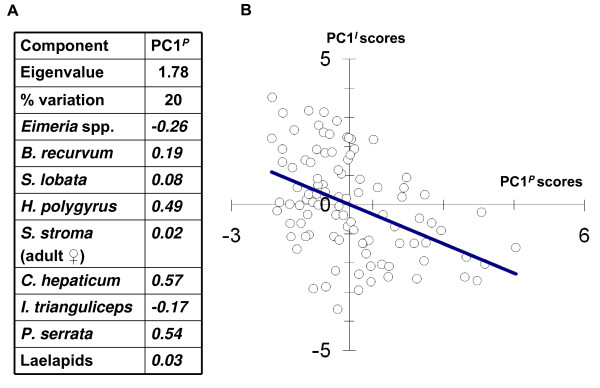
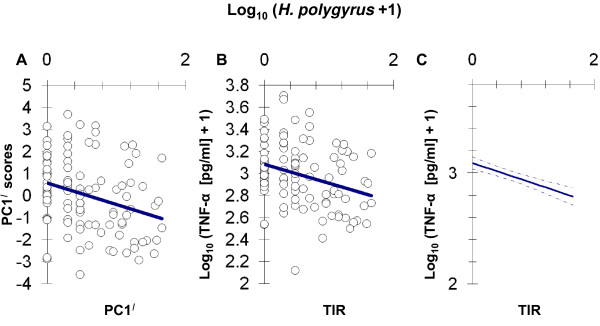
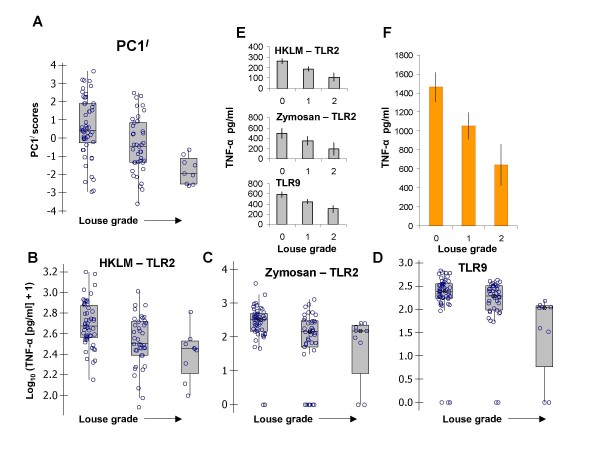
Results: Amongst a diverse group of macroparasites, only levels of the nematode Heligmosomoides polygyrus and the louse Polyplax serrata were correlated (negatively) with innate immune responsiveness (measured by splenocyte tumour necrosis factor alpha responses to a panel of toll-like receptor agonists). Polyplax serrata infection explained a strikingly high proportion of the total variation in innate responses. Contrastingly, faecal oocyst count in microparasitic Eimeria spp. was positively associated with innate immune responsiveness, most significantly for the endosomal receptors TLR7 and TLR9.
Conclusion: Analogy with relevant laboratory models suggests the underlying causality for the observed patterns may be parasite-driven immunomodulatory effects on the host. A subset of immunomodulatory parasite species could thus have a key role in structuring other infections in natural vertebrate populations by affecting the 'upstream' innate mediators, like toll-like receptors, that are important in initiating immunity. Furthermore, the magnitude of the present result suggests that populations free from immunosuppressive parasites may exist at 'unnaturally' elevated levels of innate immune activation, perhaps leading to an increased risk of immunopathology.
Figures
References
-
- Kemp AS, Mullins RJ, Weiner JM. The allergy epidemic: what is the Australian response? Med J Aust. 2006;185:226–227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
