

Cassava brown streak virus (Potyviridae) encodes a putative Maf/HAM1 pyrophosphatase implicated in reduction of mutations and a P1 proteinase that suppresses RNA silencing but contains no HC-Pro
- PMID: 19386713
- PMCID: PMC2698530
- DOI: 10.1128/JVI.00537-09
Cassava brown streak virus (Potyviridae) encodes a putative Maf/HAM1 pyrophosphatase implicated in reduction of mutations and a P1 proteinase that suppresses RNA silencing but contains no HC-Pro
Abstract
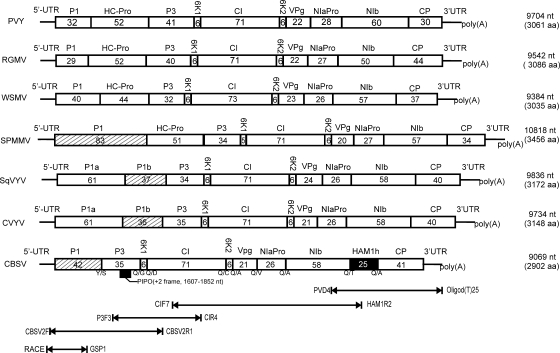
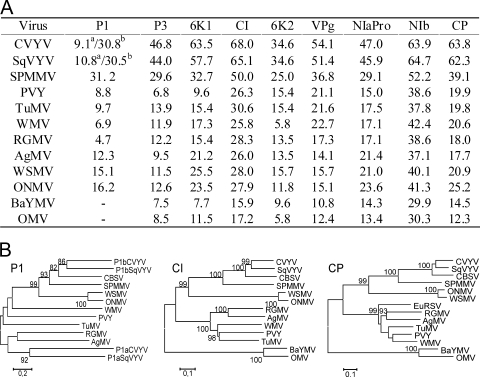
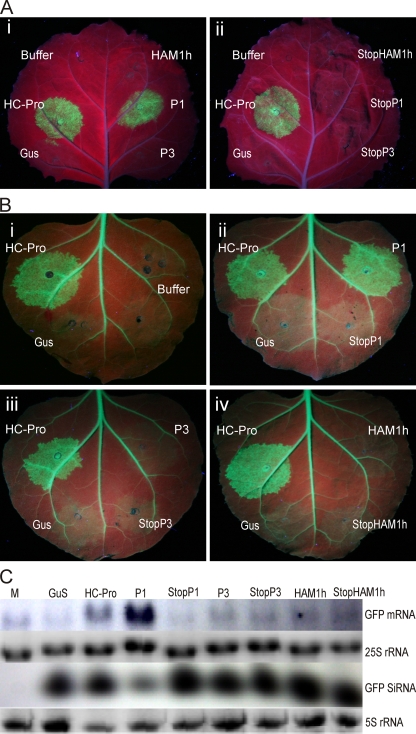
The complete positive-sense single-stranded RNA genome of Cassava brown streak virus (CBSV; genus Ipomovirus; Potyviridae) was found to consist of 9,069 nucleotides and predicted to produce a polyprotein of 2,902 amino acids. It was lacking helper-component proteinase but contained a single P1 serine proteinase that strongly suppressed RNA silencing. Besides the exceptional structure of the 5'-proximal part of the genome, CBSV also contained a Maf/HAM1-like sequence (678 nucleotides, 226 amino acids) recombined between the replicase and coat protein domains in the 3'-proximal part of the genome, which is highly conserved in Potyviridae. HAM1 was flanked by consensus proteolytic cleavage sites for ipomovirus NIaPro cysteine proteinase. Homology of CBSV HAM1 with cellular Maf/HAM1 pyrophosphatases suggests that it may intercept noncanonical nucleoside triphosphates to reduce mutagenesis of viral RNA.
Figures

Similar articles
-
Maf/ham1-like pyrophosphatases of non-canonical nucleotides are host-specific partners of viral RNA-dependent RNA polymerases.PLoS Pathog. 2022 Feb 18;18(2):e1010332. doi: 10.1371/journal.ppat.1010332. eCollection 2022 Feb. PLoS Pathog. 2022. PMID: 35180277 Free PMC article.
-
Cassava brown streak virus Ham1 protein hydrolyses mutagenic nucleotides and is a necrosis determinant.Mol Plant Pathol. 2019 Aug;20(8):1080-1092. doi: 10.1111/mpp.12813. Epub 2019 Jun 1. Mol Plant Pathol. 2019. PMID: 31154674 Free PMC article.
-
Evolution of cassava brown streak disease-associated viruses.J Gen Virol. 2011 Apr;92(Pt 4):974-87. doi: 10.1099/vir.0.026922-0. Epub 2010 Dec 15. J Gen Virol. 2011. PMID: 21169213
-
Viral inosine triphosphatase: A mysterious enzyme with typical activity, but an atypical function.Mol Plant Pathol. 2021 Mar;22(3):382-389. doi: 10.1111/mpp.13021. Epub 2021 Jan 20. Mol Plant Pathol. 2021. PMID: 33471956 Free PMC article. Review.
-
Ipomovirus--an atypical genus in the family Potyviridae transmitted by whiteflies.Pest Manag Sci. 2014 Oct;70(10):1553-67. doi: 10.1002/ps.3735. Epub 2014 Mar 3. Pest Manag Sci. 2014. PMID: 24464680 Review.
Cited by
-
Mixed infection of ITPase-encoding potyvirid and secovirid in Mercurialis perennis: evidences for a convergent euphorbia-specific viral counterstrike.Virol J. 2024 Jan 4;21(1):6. doi: 10.1186/s12985-023-02257-y. Virol J. 2024. PMID: 38178191 Free PMC article.
-
A plant virus evolved by acquiring multiple nonconserved genes to extend its host range.Proc Natl Acad Sci U S A. 2011 Oct 18;108(42):17366-71. doi: 10.1073/pnas.1113227108. Epub 2011 Oct 10. Proc Natl Acad Sci U S A. 2011. PMID: 21987809 Free PMC article.
-
Cassava brown streak disease: historical timeline, current knowledge and future prospects.Mol Plant Pathol. 2018 May;19(5):1282-1294. doi: 10.1111/mpp.12613. Epub 2017 Dec 28. Mol Plant Pathol. 2018. PMID: 28887856 Free PMC article. Review.
-
Screening for Resistance in Farmer-Preferred Cassava Cultivars from Ghana to a Mixed Infection of CBSV and UCBSV.Plants (Basel). 2020 Aug 13;9(8):1026. doi: 10.3390/plants9081026. Plants (Basel). 2020. PMID: 32823622 Free PMC article.
-
Maf/ham1-like pyrophosphatases of non-canonical nucleotides are host-specific partners of viral RNA-dependent RNA polymerases.PLoS Pathog. 2022 Feb 18;18(2):e1010332. doi: 10.1371/journal.ppat.1010332. eCollection 2022 Feb. PLoS Pathog. 2022. PMID: 35180277 Free PMC article.
References
-
- Adams, M. J., J. F. Antoniw, and F. C. Beaudoin. 2005. Overview and analysis of the polyprotein cleavage sites in the family Potyviridae. Mol. Plant Pathol. 6471-487. - PubMed
-
- Alicai, T., C. A. Omongo, M. N. Maruthi, R. J. Hillocks, Y. Baguma, R. Kawuki, A. Bua, G. W. Otim-Nape, and J. Colvin. 2007. Re-emergence of cassava brown streak disease in Uganda. Plant Dis. 9124-29. - PubMed
-
- Arazi, T., S. H. Slutsky, Y. M. Shiboleth, Y. Wang, M. Rubinstein, S. Barak, J. Yang, and A. Gal-On. 2001. Engineering zucchini yellow mosaic potyvirus as a non-pathogenic vector for expression of heterologous proteins in cucurbits. J. Biotechnol. 8767-82. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
