

Specificity of cytoplasmic dynein subunits in discrete membrane-trafficking steps
- PMID: 19386764
- PMCID: PMC2695796
- DOI: 10.1091/mbc.e08-12-1160
Specificity of cytoplasmic dynein subunits in discrete membrane-trafficking steps
Abstract
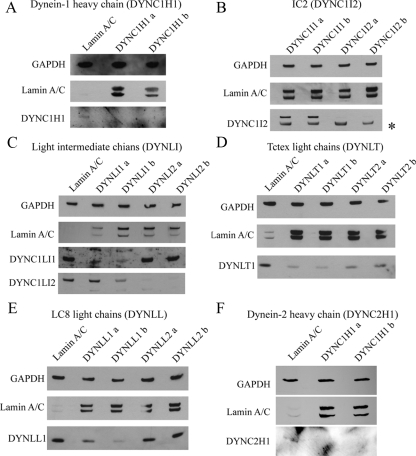
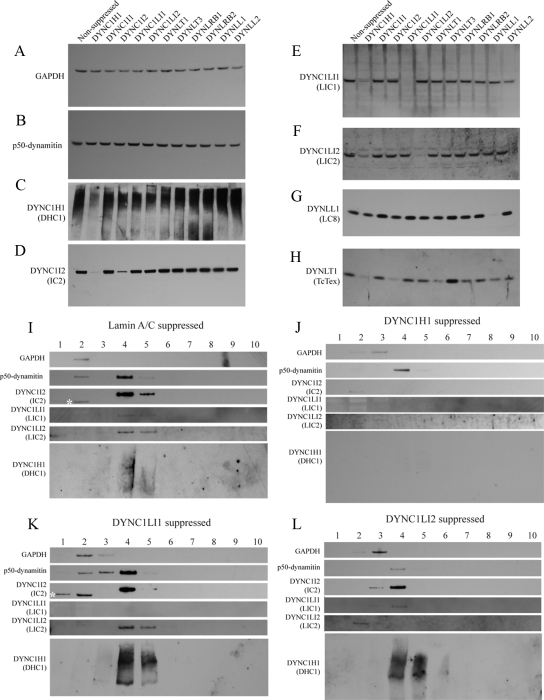
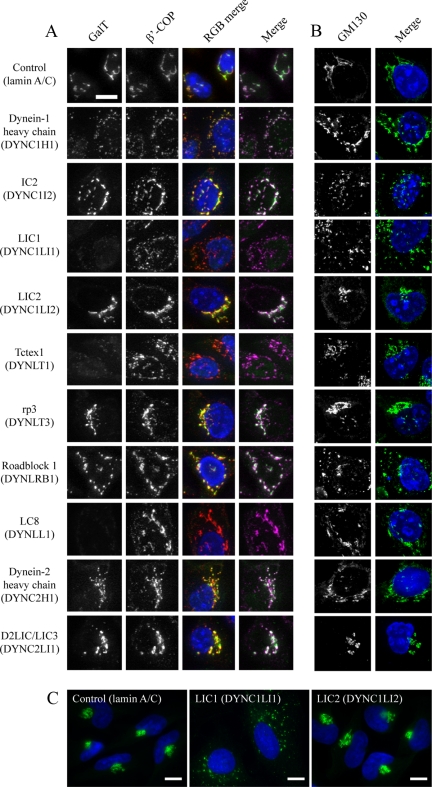
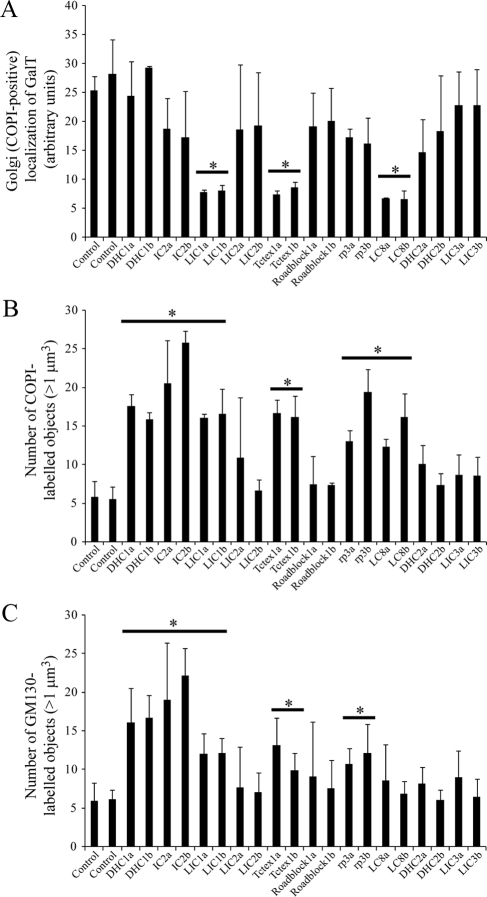
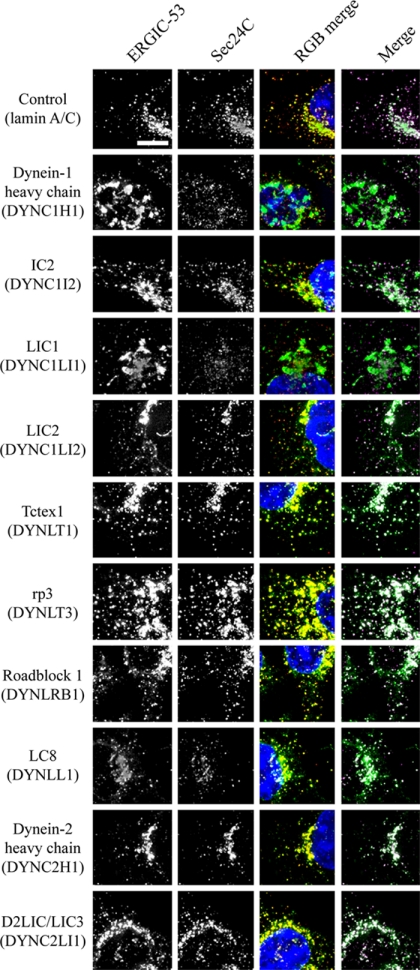
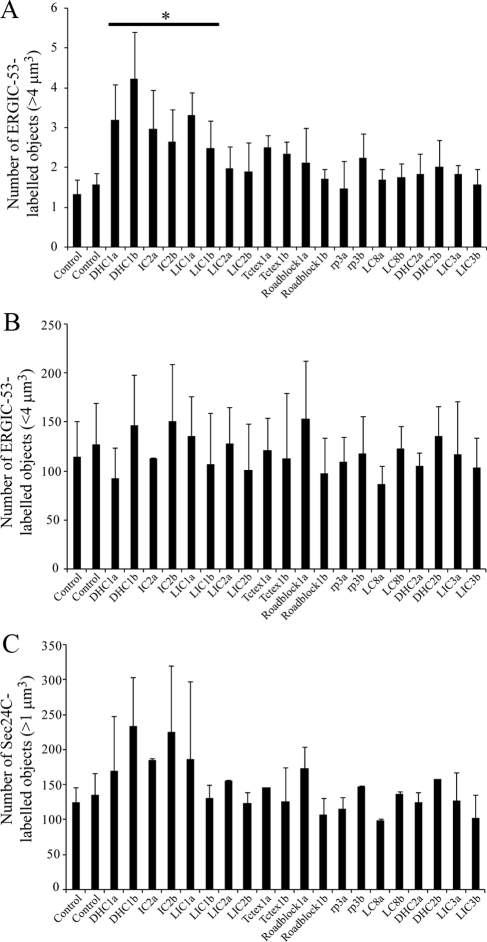
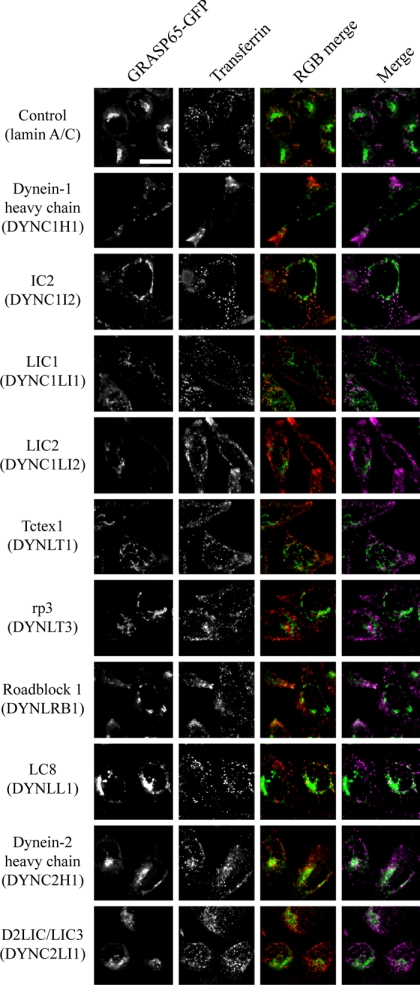
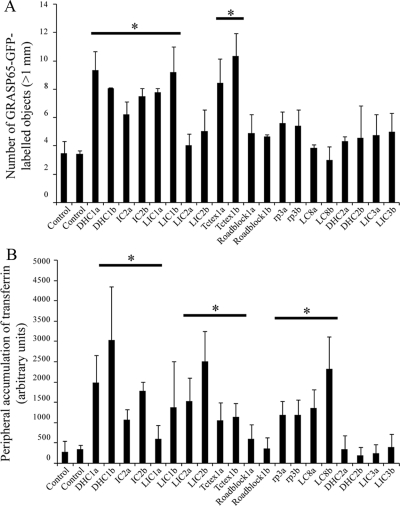
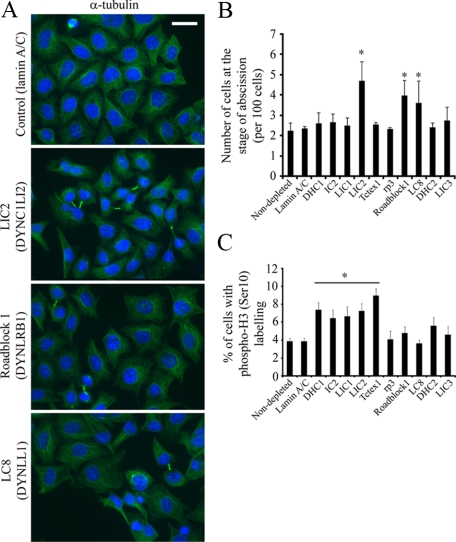
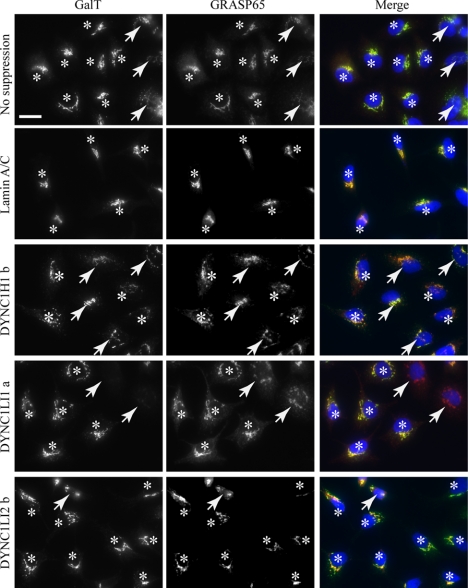
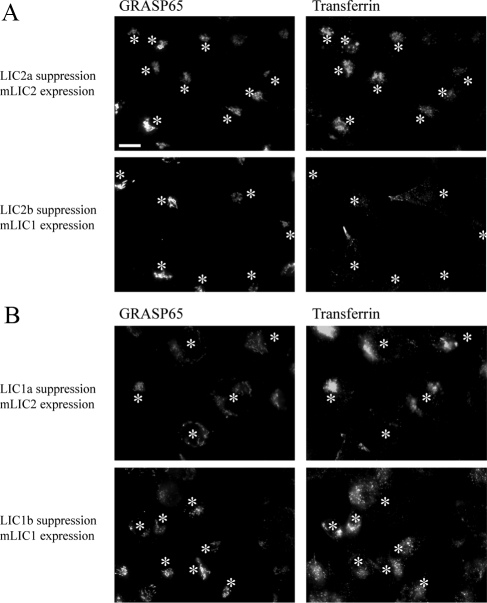
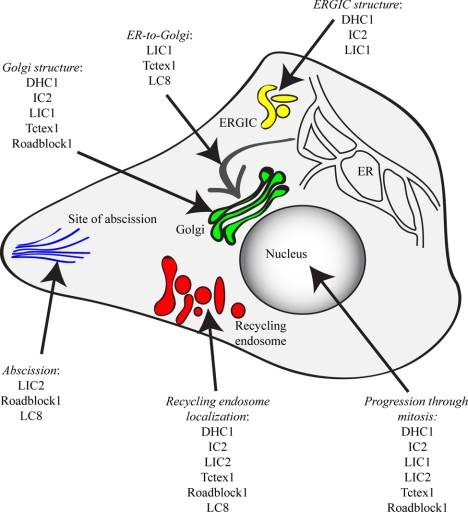
The cytoplasmic dynein motor complex is known to exist in multiple forms, but few specific functions have been assigned to individual subunits. A key limitation in the analysis of dynein in intact mammalian cells has been the reliance on gross perturbation of dynein function, e.g., inhibitory antibodies, depolymerization of the entire microtubule network, or the use of expression of dominant negative proteins that inhibit dynein indirectly. Here, we have used RNAi and automated image analysis to define roles for dynein subunits in distinct membrane-trafficking processes. Depletion of a specific subset of dynein subunits, notably LIC1 (DYNC1LI1) but not LIC2 (DYNC1LI2), recapitulates a direct block of ER export, revealing that dynein is required to maintain the steady-state composition of the Golgi, through ongoing ER-to-Golgi transport. Suppression of LIC2 but not of LIC1 results in a defect in recycling endosome distribution and cytokinesis. Biochemical analyses also define the role of each subunit in stabilization of the dynein complex; notably, suppression of DHC1 or IC2 results in concomitant loss of Tctex1. Our data demonstrate that LIC1 and LIC2 define distinct dynein complexes that function at the Golgi versus recycling endosomes, respectively, suggesting that functional populations of dynein mediate discrete intracellular trafficking pathways.
Figures
Comment in
- Mol Biol Cell. 20:2809.
References
-
- Barbar E. Dynein light chain LC8 is a dimerization hub essential in diverse protein networks. Biochemistry. 2008;47:503–508. - PubMed
-
- Benison G., Karplus P. A., Barbar E. Structure and dynamics of LC8 complexes with KXTQT-motif peptides: swallow and dynein intermediate chain compete for a common site. J. Mol. Biol. 2007;371:457–468. - PubMed
-
- Bielli A., Thornqvist P. O., Hendrick A. G., Finn R., Fitzgerald K., McCaffrey M. W. The small GTPase Rab4A interacts with the central region of cytoplasmic dynein light intermediate chain-1. Biochem. Biophys. Res. Commun. 2001;281:1141–1153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
