

Organization of intralaminar and translaminar neuronal connectivity in the superficial spinal dorsal horn
- PMID: 19386904
- PMCID: PMC2777732
- DOI: 10.1523/JNEUROSCI.6175-08.2009
Organization of intralaminar and translaminar neuronal connectivity in the superficial spinal dorsal horn
Abstract
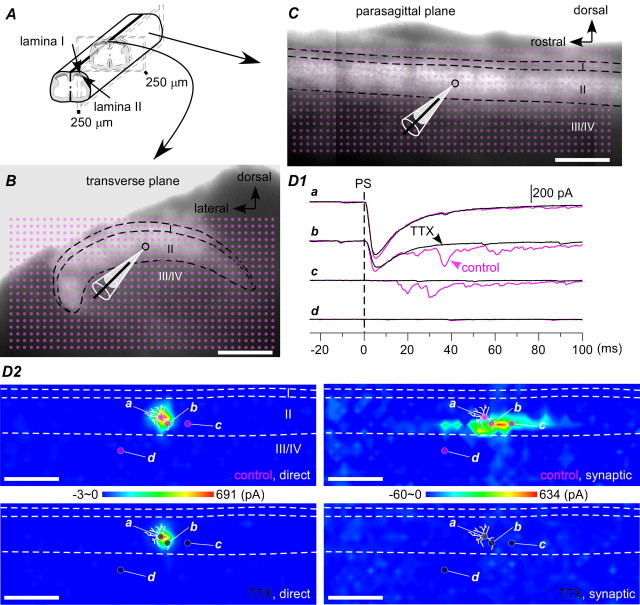
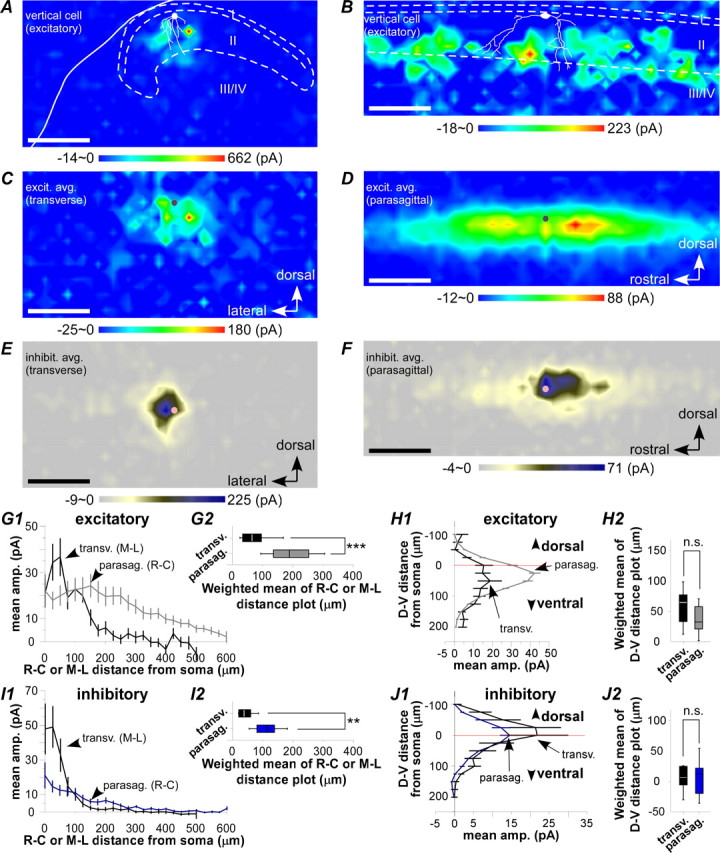
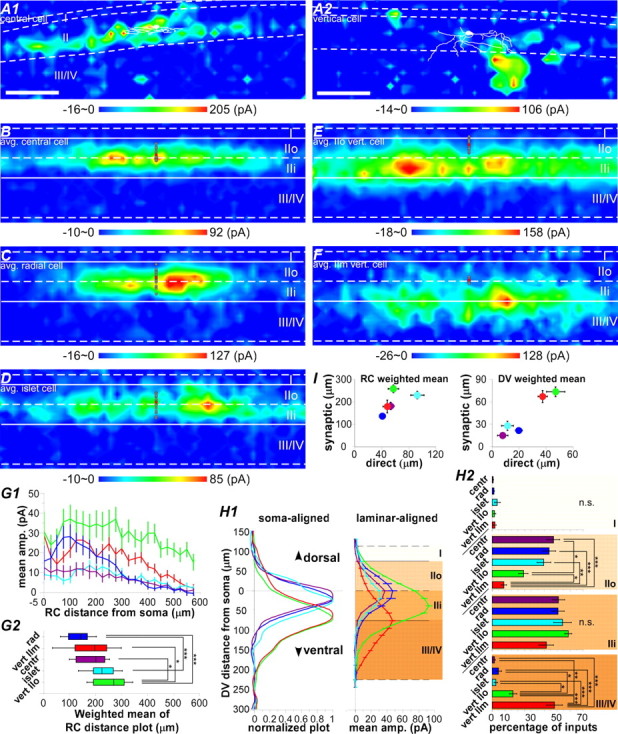
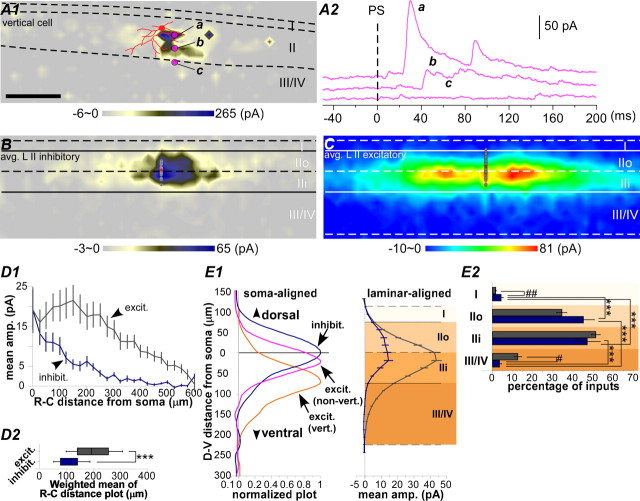
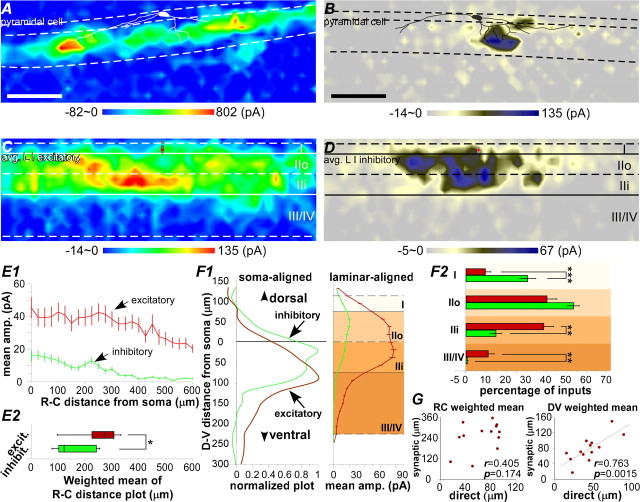
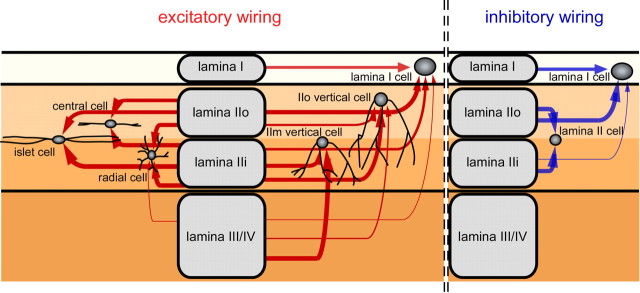
The spinal dorsal horn exhibits a high degree of intrinsic connectivity that is critical to its role in the processing of nociceptive information. To examine the spatial organization of this intrinsic connectivity, we used laser-scanning photostimulation in parasagittal and transverse slices of lumbar spinal cord to stimulate presynaptic neurons by glutamate uncaging, and mapped the location of sites that provide excitatory and inhibitory synaptic input to neurons of the superficial laminae. Excitatory interneuronal connectivity within lamina II exhibited a pronounced sagittal orientation, in keeping with the somatotopic organization present in the pattern of primary afferent projections. Excitatory inputs to all classes of lamina II neurons arose from a wider rostrocaudal area than inhibitory inputs, whereas both excitatory and inhibitory input zones were restricted mediolaterally. Lamina I-II neurons exhibited cell type-specific patterns in the laminar distribution of their excitatory inputs that were related to their dorsoventral dendritic expanse. All cell types received excitatory input predominantly from positions ventral to that of their soma, but in lamina I neurons and lamina II vertical cells this ventral displacement of the excitatory input zone was greater than in the other cell types, resulting in a more pronounced translaminar input pattern. A previously unknown excitatory input to the superficial dorsal horn from lamina III-IV was identified in a subset of the vertical cell population. These results reveal a specific three-dimensional organization in the local patterns of excitatory and inhibitory connectivity that has implications for the processing of information related to both somatotopy and sensory modality.
Figures
Similar articles
-
Three-dimensional organization of local excitatory and inhibitory inputs to neurons in laminae III-IV of the spinal dorsal horn.J Physiol. 2013 Nov 15;591(22):5645-60. doi: 10.1113/jphysiol.2013.256016. Epub 2013 Aug 27. J Physiol. 2013. PMID: 23981716 Free PMC article.
-
Subpopulation-specific patterns of intrinsic connectivity in mouse superficial dorsal horn as revealed by laser scanning photostimulation.J Physiol. 2013 Apr 1;591(7):1935-49. doi: 10.1113/jphysiol.2012.244210. Epub 2013 Jan 7. J Physiol. 2013. PMID: 23297304 Free PMC article.
-
Neurons in the lateral part of the lumbar spinal cord show distinct novel axon trajectories and are excited by short propriospinal ascending inputs.Brain Struct Funct. 2016 May;221(4):2343-60. doi: 10.1007/s00429-015-1046-3. Epub 2015 Apr 26. Brain Struct Funct. 2016. PMID: 25912439
-
Anatomy of primary afferents and projection neurones in the rat spinal dorsal horn with particular emphasis on substance P and the neurokinin 1 receptor.Exp Physiol. 2002 Mar;87(2):245-9. doi: 10.1113/eph8702351. Exp Physiol. 2002. PMID: 11856970 Review.
-
Pre- and postsynaptic inhibitory control in the spinal cord dorsal horn.Ann N Y Acad Sci. 2013 Mar;1279:90-6. doi: 10.1111/nyas.12056. Ann N Y Acad Sci. 2013. PMID: 23531006 Free PMC article. Review.
Cited by
-
The Functional Organization of Neocortical Networks Investigated in Slices with Local Field Recordings and Laser Scanning Photostimulation.PLoS One. 2015 Jul 2;10(7):e0132008. doi: 10.1371/journal.pone.0132008. eCollection 2015. PLoS One. 2015. PMID: 26134668 Free PMC article.
-
The search for novel analgesics: re-examining spinal cord circuits with new tools.Front Pharmacol. 2014 Feb 25;5:22. doi: 10.3389/fphar.2014.00022. eCollection 2014. Front Pharmacol. 2014. PMID: 24616699 Free PMC article.
-
Peptidergic CGRPα primary sensory neurons encode heat and itch and tonically suppress sensitivity to cold.Neuron. 2013 Apr 10;78(1):138-51. doi: 10.1016/j.neuron.2013.01.030. Epub 2013 Mar 21. Neuron. 2013. PMID: 23523592 Free PMC article.
-
Monosynaptic excitatory inputs to spinal lamina I anterolateral-tract-projecting neurons from neighbouring lamina I neurons.J Physiol. 2010 Nov 15;588(Pt 22):4489-505. doi: 10.1113/jphysiol.2010.197012. Epub 2010 Sep 27. J Physiol. 2010. PMID: 20876196 Free PMC article.
-
Lesion of the dopaminergic nigrostriatal pathway induces trigeminal dynamic mechanical allodynia.Brain Behav. 2014 May;4(3):368-80. doi: 10.1002/brb3.214. Epub 2014 Feb 20. Brain Behav. 2014. PMID: 24944866 Free PMC article.
References
-
- Baba H, Ji RR, Kohno T, Moore KA, Ataka T, Wakai A, Okamoto M, Woolf CJ. Removal of GABAergic inhibition facilitates polysynaptic A fiber-mediated excitatory transmission to the superficial spinal dorsal horn. Mol Cell Neurosci. 2003;24:818–830. - PubMed
-
- Bentley GN, Gent JP. Electrophysiological properties of substantia gelatinosa neurones in a novel adult spinal slice preparation. J Neurosci Methods. 1994;53:157–162. - PubMed
-
- Braz JM, Nassar MA, Wood JN, Basbaum AI. Parallel “pain” pathways arise from subpopulations of primary afferent nociceptor. Neuron. 2005;47:787–793. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases