

Functional analysis of -351 interleukin-9 promoter polymorphism reveals an activator controlled by NF-kappaB
- PMID: 19387455
- PMCID: PMC2702712
- DOI: 10.1038/gene.2009.28
Functional analysis of -351 interleukin-9 promoter polymorphism reveals an activator controlled by NF-kappaB
Abstract
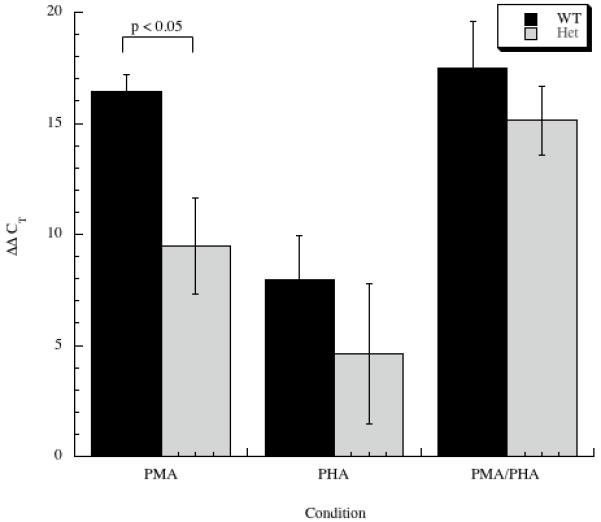
Genetic studies have shown linkages for asthma to the chromosomal region 5q31-q33 in humans that includes the IL-9 gene. An A-to-G base substitution has been identified at bp -351 in the IL-9 promoter. The role of this polymorphism in IL-9 promoter function was assessed utilizing CD4+ T cells purified from individuals with one or two of the G alleles in comparison to those homozygous for the wild-type A. The presence of an A at -351 (A allele) increased mitogen-stimulated IL-9 transcription twofold in comparison to subjects with one or two G alleles at this position. Binding of nuclear extract proteins from IL-9-producing human cell lines to DNA sequences including this base exchange demonstrated specific binding of the transcription factor NF-kappaB. Binding of NF-kappaB to the IL-9 promoter was confirmed in vivo using the chromatin immunoprecipitation assay. Recombinant NF-kappaB bound to a promoter fragment with the A allele with fivefold higher affinity than it did to a promoter with the G allele. Individuals carrying the A allele of the IL-9 promoter display increased synthesis of IL-9, which may result in strong Th2 immune responses and a modulation of their susceptibility to infectious, neoplastic, parasitic or atopic disease.
Figures








References
-
- Borish LC, Steinke JW. Cytokines and chemokines. J Allergy Clin Immunol. 2003;111:S460–475. - PubMed
-
- Hultner L, Moeller J. Mast cell growth-enhancing activity (MEA) stimulates interleukin 6 production in a mouse bone marrow-derived mast cell line and a malignant subline. Exp Hematol. 1990;18:873–877. - PubMed
-
- Renauld J-C, Kermouni A, Vink A, Louahed J, Van Snick J. Interleukin-9 and its receptor: involvement in mast cell differentiation and T cell oncogenesis. J Leukoc Biol. 1995;57:353–360. - PubMed
-
- Louahed J, Kermouni A, Van Snick J, Renauld J-C. IL-9 induces expression of granzymes and high-affinity IgE receptor in murine T helper clones. J Immunol. 1995;154:5061–5070. - PubMed
-
- Hultner L, Druez C, Moeller J, et al. Mast cell growth-enhancing activity (MEA) is structurally related and functionally identical to the novel mouse T cell growth factor P40/TCGFIII (interleukin 9) Eur J Immunol. 1990;20:1413–1416. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
