

Deletion of (54)FLRAPSWF(61) residues decreases the oligomeric size and enhances the chaperone function of alphaB-crystallin
- PMID: 19388699
- PMCID: PMC3997080
- DOI: 10.1021/bi900085v
Deletion of (54)FLRAPSWF(61) residues decreases the oligomeric size and enhances the chaperone function of alphaB-crystallin
Abstract
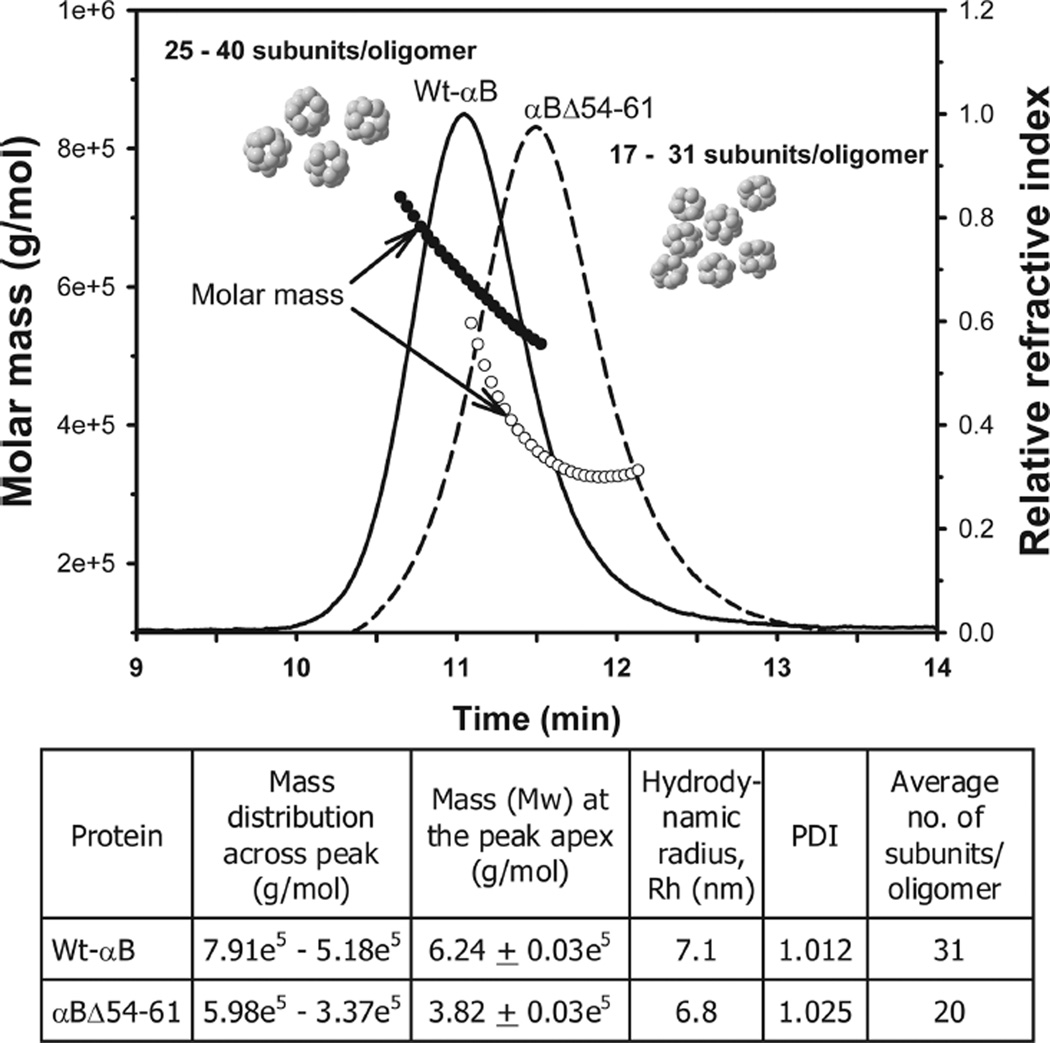
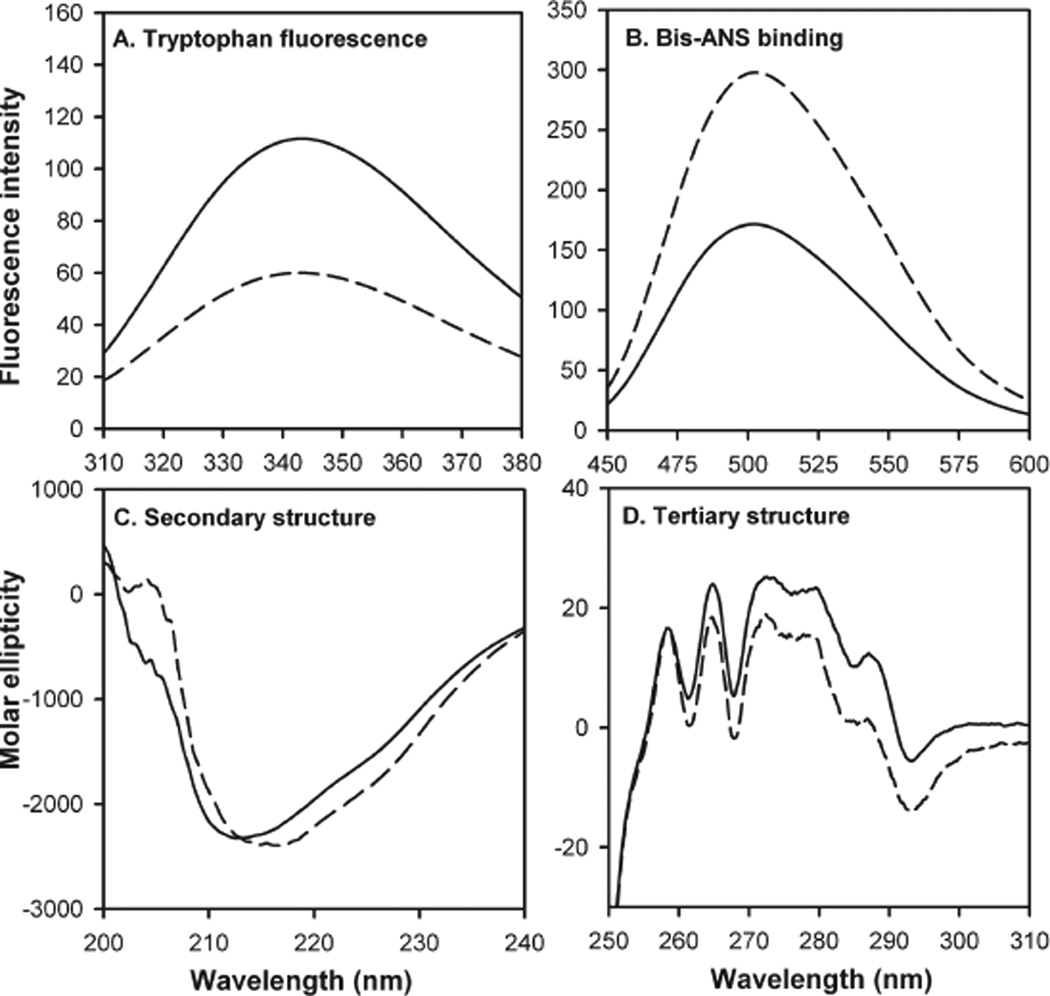
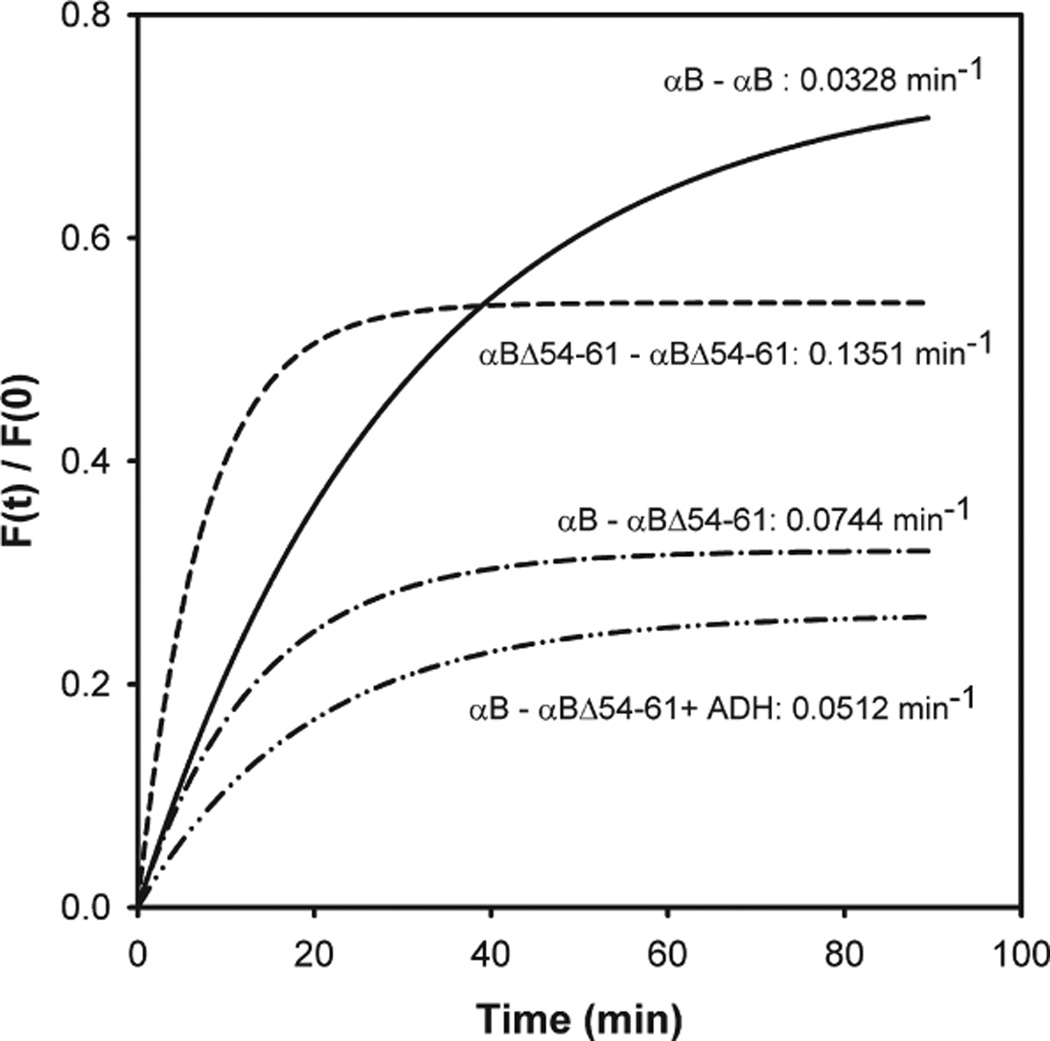
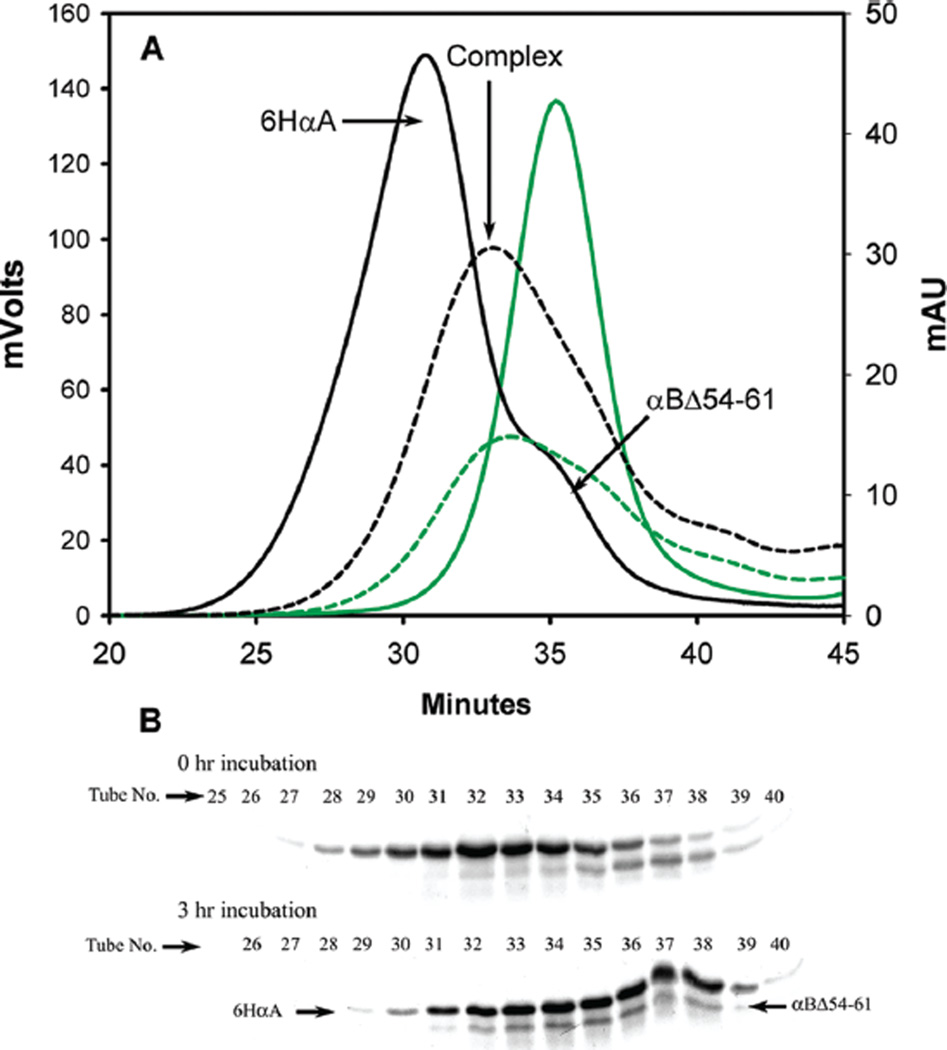
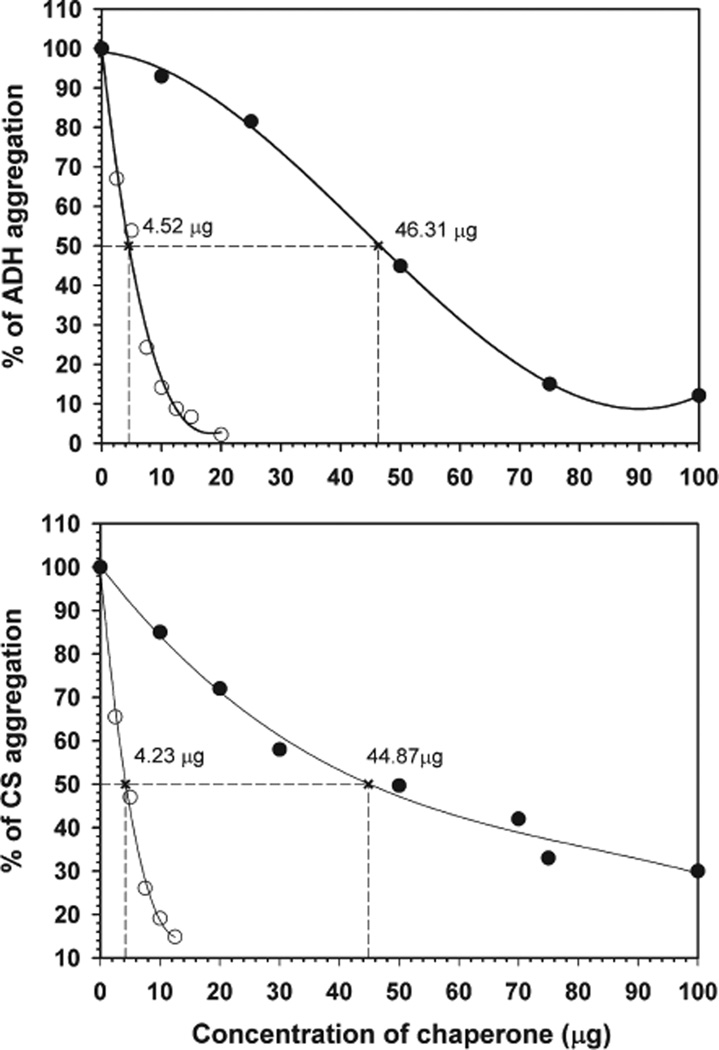
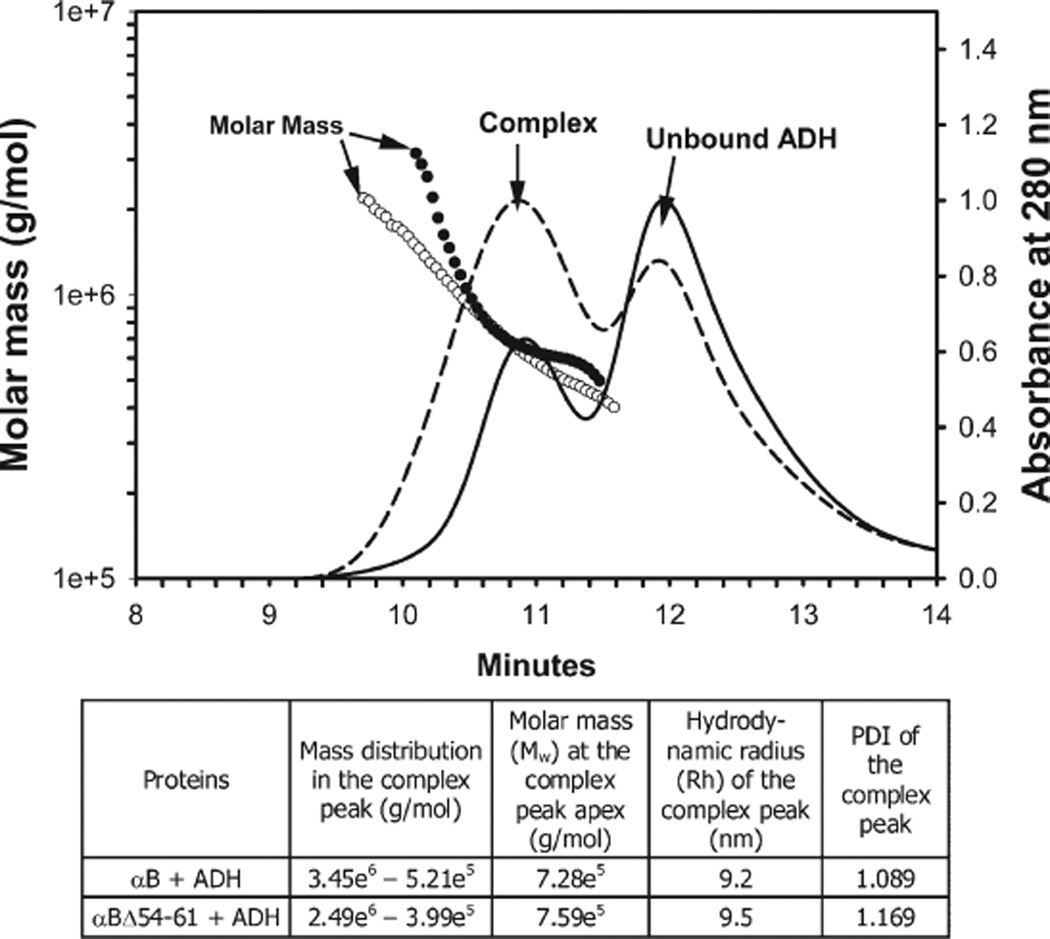
AlphaB-crystallin is a member of the small heat shock protein family and is known to have chaperone activity. Using a peptide scan approach, we previously determined that regions 42-57, 60-71, and 88-123 in alphaB-crystallin interact with alphaA-crystallin during heterooligomer formation. To further characterize the significance of the N-terminal domain of alphaB-crystallin, we prepared a deletion mutant that lacks residues (54)FLRAPSWF(61) (alphaBDelta54-61) and found that the absence of residues 54-61 in alphaB-crystallin significantly decreased the homooligomeric mass of alphaB-crystallin. The average oligomeric mass of wild-type alphaB-crystallin and of alphaBDelta54-61, calculated using multiangle light scattering, was 624 and 382 kDa, respectively. The mutant subunits aggregate to form smaller, less-compact oligomers with a 4-fold increase in subunit exchange rate. Deletion of the 54-61 region resulted in a 50% decrease in intrinsic tryptophan fluorescence. The alphaBDelta54-61 mutant showed a 2-fold increase in 1,1'-bi(4-anilino)naphthalene-5,5'-disulfonic acid (bis-ANS) binding as compared to the wild-type protein, suggesting increased hydrophobicity of the mutant protein. Accompanying the evidence of increased hydrophobicity in the deletion mutant was a 10-fold increase in antiaggregation activity. Homooligomers of 6HalphaA (750 kDa) readily exchanged subunits with alphaBDelta54-61 homooligomers at 37 degrees C, forming heterooligomers with an intermediate mass of 625 kDa. Our data suggest that residues (54)FLRAPSWF(61) contribute to the higher order assembly of alphaB-crystallin oligomers. Residues (54)FLRAPSWF(61) in alphaB-crystallin are not essential for target protein binding during chaperone action, but this region apparently has a role in the chaperone activity of native alphaB-crystallin.
Figures


Similar articles
-
AlphaA-crystallin interacting regions in the small heat shock protein, alphaB-crystallin.Biochemistry. 2004 Dec 21;43(50):15785-95. doi: 10.1021/bi048151s. Biochemistry. 2004. PMID: 15595834
-
The interaction between alphaA- and alphaB-crystallin is sequence-specific.Mol Vis. 2006 May 24;12:581-7. Mol Vis. 2006. PMID: 16760894
-
Structural and functional roles of deamidation and/or truncation of N- or C-termini in human alpha A-crystallin.Biochemistry. 2008 Sep 23;47(38):10069-83. doi: 10.1021/bi8001902. Epub 2008 Aug 29. Biochemistry. 2008. PMID: 18754677
-
Regulation of αA- and αB-crystallins via phosphorylation in cellular homeostasis.Cell Mol Life Sci. 2015 Nov;72(21):4127-37. doi: 10.1007/s00018-015-1996-x. Epub 2015 Jul 26. Cell Mol Life Sci. 2015. PMID: 26210153 Free PMC article. Review.
-
One size does not fit all: the oligomeric states of αB crystallin.FEBS Lett. 2013 Apr 17;587(8):1073-80. doi: 10.1016/j.febslet.2013.01.021. Epub 2013 Jan 20. FEBS Lett. 2013. PMID: 23340341 Free PMC article. Review.
Cited by
-
Structural and functional consequences of chaperone site deletion in αA-crystallin.Biochim Biophys Acta. 2016 Nov;1864(11):1529-38. doi: 10.1016/j.bbapap.2016.08.006. Epub 2016 Aug 11. Biochim Biophys Acta. 2016. PMID: 27524665 Free PMC article.
-
Acetylation of αA-crystallin in the human lens: effects on structure and chaperone function.Biochim Biophys Acta. 2012 Feb;1822(2):120-9. doi: 10.1016/j.bbadis.2011.11.011. Epub 2011 Nov 18. Biochim Biophys Acta. 2012. PMID: 22120592 Free PMC article.
-
Proteinaceous Transformers: Structural and Functional Variability of Human sHsps.Int J Mol Sci. 2020 Jul 30;21(15):5448. doi: 10.3390/ijms21155448. Int J Mol Sci. 2020. PMID: 32751672 Free PMC article. Review.
-
Succinylation Is a Gain-of-Function Modification in Human Lens αB-Crystallin.Biochemistry. 2019 Mar 5;58(9):1260-1274. doi: 10.1021/acs.biochem.8b01053. Epub 2019 Feb 20. Biochemistry. 2019. PMID: 30758948 Free PMC article.
-
Effect of Structural Changes Induced by Deletion of 54FLRAPSWF61 Sequence in αB-crystallin on Chaperone Function and Anti-Apoptotic Activity.Int J Mol Sci. 2021 Oct 5;22(19):10771. doi: 10.3390/ijms221910771. Int J Mol Sci. 2021. PMID: 34639110 Free PMC article.
References
-
- Horwitz J. Alpha-crystallin. Exp. Eye Res. 2003;76:145–153. - PubMed
-
- Bhat SP, Nagineni CN. alpha B subunit of lens-specific protein alpha-crystallin is present in other ocular and non-ocular tissues. Biochem. Biophys. Res. Commun. 1989;158:319–325. - PubMed
-
- Clark JI, Muchowski PJ. Small heat-shock proteins and their potential role in human disease. Curr. Opin. Struct. Biol. 2000;10:52–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
