

Sizing up ethanol-induced plasticity: the role of small and large conductance calcium-activated potassium channels
- PMID: 19389201
- PMCID: PMC2760381
- DOI: 10.1111/j.1530-0277.2009.00936.x
Sizing up ethanol-induced plasticity: the role of small and large conductance calcium-activated potassium channels
Abstract
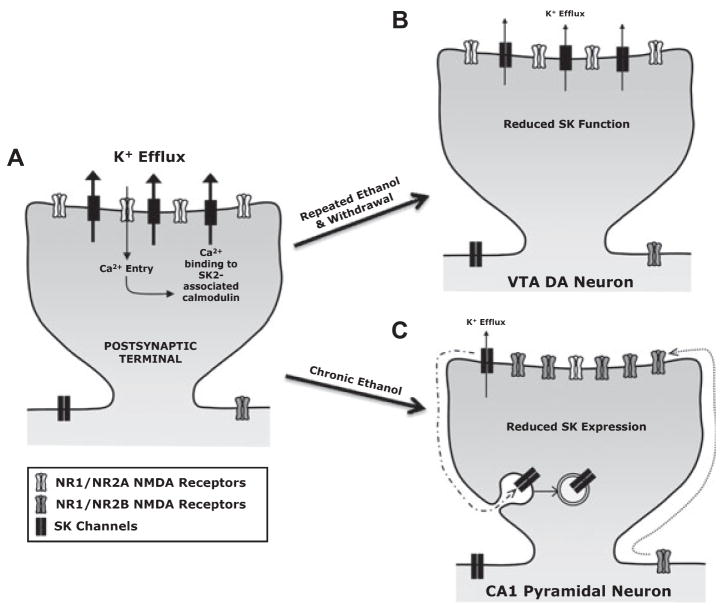
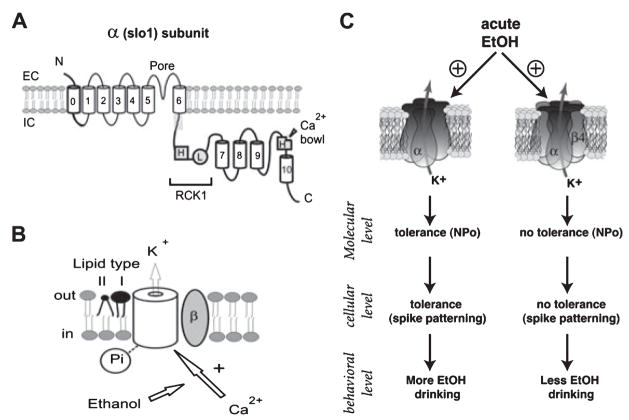
Small (SK) and large conductance (BK) Ca(2+)-activated K(+) channels contribute to action potential repolarization, shape dendritic Ca(2+)spikes and postsynaptic responses, modulate the release of hormones and neurotransmitters, and contribute to hippocampal-dependent synaptic plasticity. Over the last decade, SK and BK channels have emerged as important targets for the development of acute ethanol tolerance and for altering neuronal excitability following chronic ethanol consumption. In this mini-review, we discuss new evidence implicating SK and BK channels in ethanol tolerance and ethanol-associated homeostatic plasticity. Findings from recent reports demonstrate that chronic ethanol produces a reduction in the function of SK channels in VTA dopaminergic and CA1 pyramidal neurons. It is hypothesized that the reduction in SK channel function increases the propensity for burst firing in VTA neurons and increases the likelihood for aberrant hyperexcitability during ethanol withdrawal in hippocampus. There is also increasing evidence supporting the idea that ethanol sensitivity of native BK channel results from differences in BK subunit composition, the proteolipid microenvironment, and molecular determinants of the channel-forming subunit itself. Moreover, these molecular entities play a substantial role in controlling the temporal component of ethanol-associated neuroadaptations in BK channels. Taken together, these studies suggest that SK and BK channels contribute to ethanol tolerance and adaptive plasticity.
Figures
References
-
- Anderson NJ, Slough S, Watson WP. In vivo characterisation of the small-conductance KCa (SK) channel activator 1-ethyl-2-benzimidazolinone (1-EBIO) as a potential anticonvulsant. Eur J Pharmacol. 2006;546:48–53. - PubMed
-
- Bailey CP, Manley SJ, Watson WP, Wonnacott S, Molleman A, Little HJ. Chronic ethanol administration alters activity in ventral tegmental area neurons after cessation of withdrawal hyperexcitability. Brain Res. 1998;803:144–152. - PubMed
-
- Bailey CP, O’Callaghan MJ, Croft AP, Manley SJ, Little HJ. Alterations in mesolimbic dopamine function during the abstinence period following chronic ethanol consumption. Neuropharmacology. 2001;41:989–999. - PubMed
-
- Bond CT, Maylie J, Adelman JP. SK channels in excitability, pacemaking and synaptic integration. Curr Opin Neurobiol. 2005;15:305–311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
