

Jagged 1 is necessary for normal mouse lens formation
- PMID: 19389370
- PMCID: PMC2674137
- DOI: 10.1016/j.ydbio.2009.01.015
Jagged 1 is necessary for normal mouse lens formation
Abstract
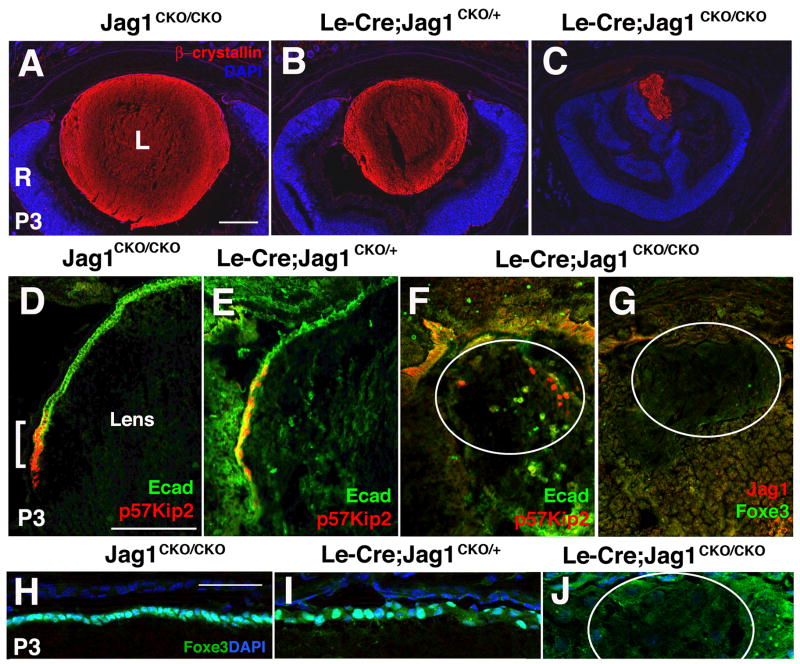
In mammals, two spatially and temporally distinct waves of fiber cell differentiation are crucial steps for normal lens development. In between these phases, an anterior growth zone forms in which progenitor cells migrate circumferentially, terminally exit the cell cycle and initiate differentiation at the lens equator. Much remains unknown about the molecular pathways orchestrating these processes. Previously, the Notch signal transduction pathway was shown to be critical for anterior lens progenitor cell growth and differentiation. However, the ligand or ligand(s) that direct these events are unknown. Using conditional gene targeting, we show that Jagged1 is required for lens fiber cell genesis, particularly that of secondary fiber cells. In the absence of Jagged1, the anterior growth and equatorial transition zones fail to develop fully, with only a handful of differentiated fiber cells present at birth. Adult Jagged1 conditional mutants completely lack lenses, along with severe anterior chamber deformities. Our data support the hypothesis that Jagged1-Notch signaling conveys a lateral inductive signal, which is indispensable for lens progenitor cell proliferation and differentiation.
Figures





Similar articles
-
Requirements for Jag1-Rbpj mediated Notch signaling during early mouse lens development.Dev Dyn. 2012 Mar;241(3):493-504. doi: 10.1002/dvdy.23739. Epub 2012 Jan 25. Dev Dyn. 2012. PMID: 22275127 Free PMC article.
-
Notch signaling is required for lateral induction of Jagged1 during FGF-induced lens fiber differentiation.Dev Biol. 2009 Aug 1;332(1):166-76. doi: 10.1016/j.ydbio.2009.05.566. Epub 2009 May 27. Dev Biol. 2009. PMID: 19481073 Free PMC article.
-
Jagged1 is the major regulator of Notch-dependent cell fate in proximal airways.Dev Dyn. 2013 Jun;242(6):678-86. doi: 10.1002/dvdy.23965. Epub 2013 Apr 29. Dev Dyn. 2013. PMID: 23526493
-
Functional role of Notch signaling in the developing and postnatal heart.J Mol Cell Cardiol. 2008 Oct;45(4):495-504. doi: 10.1016/j.yjmcc.2008.02.273. Epub 2008 Mar 10. J Mol Cell Cardiol. 2008. PMID: 18410944 Review.
-
Blockade of Jagged/Notch pathway abrogates transforming growth factor β2-induced epithelial-mesenchymal transition in human retinal pigment epithelium cells.Curr Mol Med. 2014 May;14(4):523-34. doi: 10.2174/1566524014666140331230411. Curr Mol Med. 2014. PMID: 24694299 Review.
Cited by
-
Multiomics Analysis Reveals Novel Genetic Determinants for Lens Differentiation, Structure, and Transparency.Biomolecules. 2023 Apr 19;13(4):693. doi: 10.3390/biom13040693. Biomolecules. 2023. PMID: 37189439 Free PMC article. Review.
-
The lens growth process.Prog Retin Eye Res. 2017 Sep;60:181-200. doi: 10.1016/j.preteyeres.2017.04.001. Epub 2017 Apr 11. Prog Retin Eye Res. 2017. PMID: 28411123 Free PMC article. Review.
-
Fibrosis in the lens. Sprouty regulation of TGFβ-signaling prevents lens EMT leading to cataract.Exp Eye Res. 2016 Jan;142:92-101. doi: 10.1016/j.exer.2015.02.004. Epub 2015 May 21. Exp Eye Res. 2016. PMID: 26003864 Free PMC article. Review.
-
The lens: a classical model of embryonic induction providing new insights into cell determination in early development.Philos Trans R Soc Lond B Biol Sci. 2011 Apr 27;366(1568):1193-203. doi: 10.1098/rstb.2010.0175. Philos Trans R Soc Lond B Biol Sci. 2011. PMID: 21402580 Free PMC article. Review.
-
Endothelial follistatin-like-1 regulates the postnatal development of the pulmonary vasculature by modulating BMP/Smad signaling.Pulm Circ. 2017 Mar 15;7(1):219-231. doi: 10.1177/2045893217702340. eCollection 2017 Mar. Pulm Circ. 2017. PMID: 28680581 Free PMC article.
References
-
- Beebe D, Garcia C, Wang X, Rajagopal R, Feldmeier M, Kim JY, Chytil A, Moses H, Ashery-Padan R, Rauchman M. Contributions by members of the TGFbeta superfamily to lens development. Int J Dev Biol. 2004;48:845–56. - PubMed
-
- Blixt A, Landgren H, Johansson BR, Carlsson P. Foxe3 is required for morphogenesis and differentiation of the anterior segment of the eye and is sensitive to Pax6 gene dosage. Dev Biol. 2007;302:218–29. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
