

Repression of Runx2 by androgen receptor (AR) in osteoblasts and prostate cancer cells: AR binds Runx2 and abrogates its recruitment to DNA
- PMID: 19389811
- PMCID: PMC2718746
- DOI: 10.1210/me.2008-0470
Repression of Runx2 by androgen receptor (AR) in osteoblasts and prostate cancer cells: AR binds Runx2 and abrogates its recruitment to DNA
Abstract
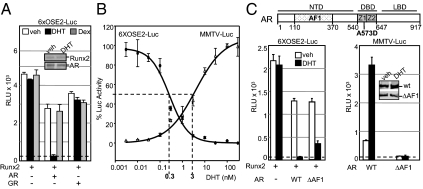
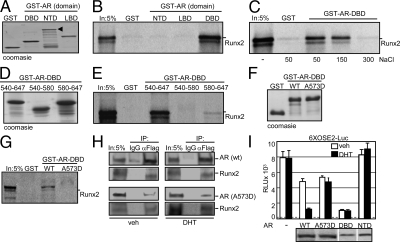
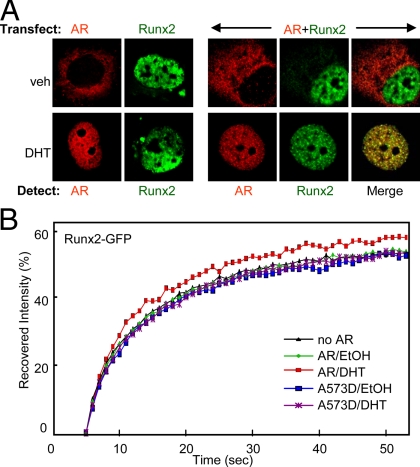
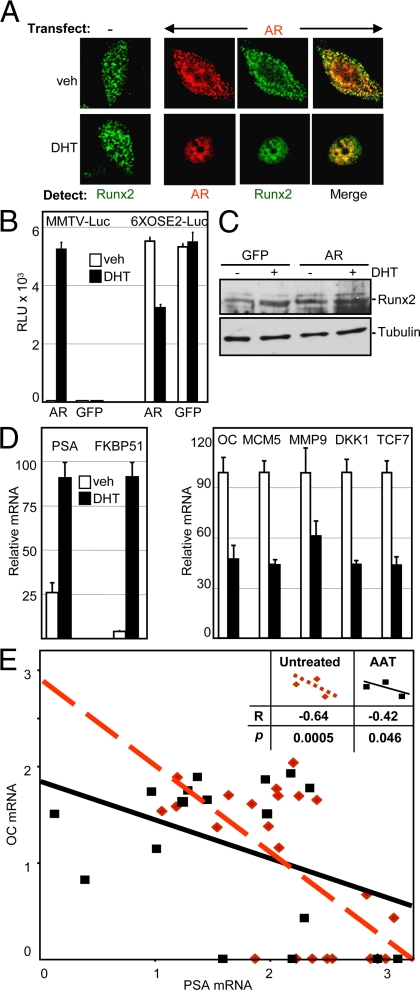
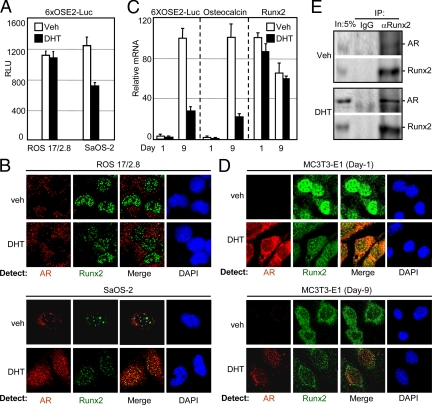
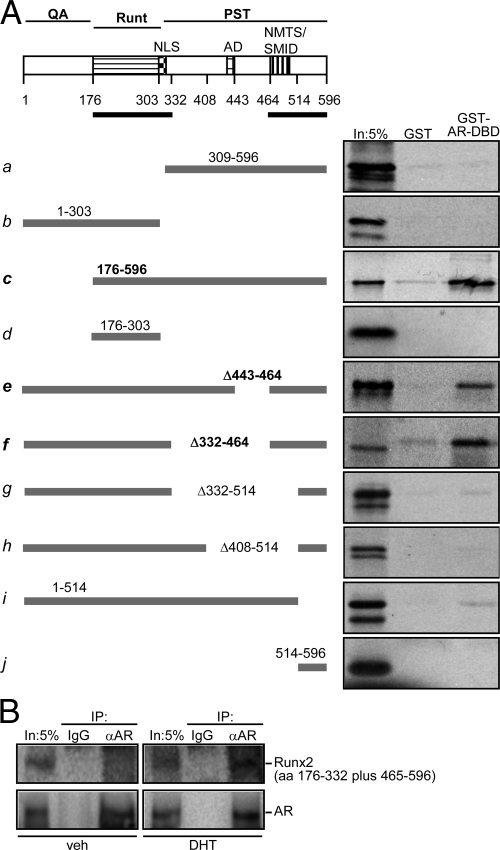
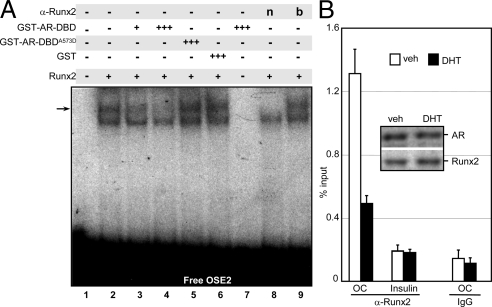
Runx2 and androgen receptor (AR) are master transcription factors with pivotal roles in bone metabolism and prostate cancer (PCa). We dissected AR-mediated repression of Runx2 in dihydrotestosterone (DHT)-treated osteoblastic and PCa cells using reporter assays and endogenous Runx2 target genes. Repression required DHT, but not AR's transactivation function, and was associated with nuclear colocalization of the two proteins. Runx2 and AR coimmunoprecipitated and interacted directly in glutathione-S-transferase pull-down assays. Interaction was ionic in nature. Intact AR DNA-binding domain (DBD) was necessary and sufficient for both interaction with Runx2 and its repression. Runx2 sequences required for interaction were the C-terminal 132 amino acid residues together with the Runt DBD. Runx2 DNA binding was abrogated by endogenous AR in chromatin immunoprecipitation assays and by recombinant AR-DBD in gel shift assays. Furthermore, AR caused increased nuclear mobility of Runx2 as indicated by faster fluorescence recovery after photobleaching. Thus, AR binds Runx2 and abrogates its binding to DNA and possibly to other nuclear components. Clinical relevance of our results was suggested by an inverse correlation between expression of AR-responsive prostate-specific antigen and osteocalcin genes in PCa biopsies. Given the tumor suppressor properties of Runx2, its repression by AR may constitute a mechanism of hormone carcinogenesis. Attenuation of Runx2 by AR in osteoblasts may play a role in skeletal metabolism: the bone-sparing effect of androgens is attributable, in part, to keeping Runx2 activity in check and preventing high-turnover bone disease such as seen after castration and in transgenic mice overexpressing Runx2 in osteoblasts.
Figures
Similar articles
-
Differential effects of RUNX2 on the androgen receptor in prostate cancer: synergistic stimulation of a gene set exemplified by SNAI2 and subsequent invasiveness.Cancer Res. 2014 May 15;74(10):2857-68. doi: 10.1158/0008-5472.CAN-13-2003. Epub 2014 Mar 19. Cancer Res. 2014. PMID: 24648349 Free PMC article.
-
Bone Cell Activity in Clinical Prostate Cancer Bone Metastasis and Its Inverse Relation to Tumor Cell Androgen Receptor Activity.Int J Mol Sci. 2018 Apr 18;19(4):1223. doi: 10.3390/ijms19041223. Int J Mol Sci. 2018. PMID: 29670000 Free PMC article.
-
General transcription factor IIA-gamma increases osteoblast-specific osteocalcin gene expression via activating transcription factor 4 and runt-related transcription factor 2.J Biol Chem. 2008 Feb 29;283(9):5542-53. doi: 10.1074/jbc.M705653200. Epub 2008 Jan 2. J Biol Chem. 2008. PMID: 18171674 Free PMC article.
-
Functional localization and competition between the androgen receptor and T-cell factor for nuclear beta-catenin: a means for inhibition of the Tcf signaling axis.Oncogene. 2003 Aug 28;22(36):5602-13. doi: 10.1038/sj.onc.1206802. Oncogene. 2003. PMID: 12944908
-
Androgen receptors in hormone-dependent and castration-resistant prostate cancer.Pharmacol Ther. 2013 Dec;140(3):223-38. doi: 10.1016/j.pharmthera.2013.07.003. Epub 2013 Jul 13. Pharmacol Ther. 2013. PMID: 23859952 Review.
Cited by
-
Integrin αvβ3 and CD44 pathways in metastatic prostate cancer cells support osteoclastogenesis via a Runx2/Smad 5/receptor activator of NF-κB ligand signaling axis.Mol Cancer. 2012 Sep 11;11:66. doi: 10.1186/1476-4598-11-66. Mol Cancer. 2012. PMID: 22966907 Free PMC article.
-
Relationship between RUNX1 and AXIN1 in ER-negative versus ER-positive Breast Cancer.Cell Cycle. 2017 Feb 16;16(4):312-318. doi: 10.1080/15384101.2016.1237325. Epub 2017 Jan 5. Cell Cycle. 2017. PMID: 28055379 Free PMC article.
-
Association of polyalanine and polyglutamine coiled coils mediates expansion disease-related protein aggregation and dysfunction.Hum Mol Genet. 2014 Jul 1;23(13):3402-20. doi: 10.1093/hmg/ddu049. Epub 2014 Feb 4. Hum Mol Genet. 2014. PMID: 24497578 Free PMC article.
-
Characterization of CD44 intracellular domain interaction with RUNX2 in PC3 human prostate cancer cells.Cell Commun Signal. 2019 Jul 22;17(1):80. doi: 10.1186/s12964-019-0395-6. Cell Commun Signal. 2019. PMID: 31331331 Free PMC article.
-
Estrogens and selective estrogen receptor modulators differentially antagonize Runx2 in ST2 mesenchymal progenitor cells.J Steroid Biochem Mol Biol. 2018 Oct;183:10-17. doi: 10.1016/j.jsbmb.2018.05.002. Epub 2018 May 8. J Steroid Biochem Mol Biol. 2018. PMID: 29751107 Free PMC article.
References
-
- Komori T, Yagi H, Nomura S, Yamaguchi A, Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, Sato M, Okamoto R, Kitamura Y, Yoshiki S, Kishimoto T 1997 Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 89:755–764 - PubMed
-
- Otto F, Thornell AP, Crompton T, Denzel A, Gilmour KC, Rosewell IR, Stamp GW, Beddington RS, Mundlos S, Olsen BR, Selby PB, Owen MJ 1997 Cbfa1, a candidate gene for cleidocranial dysplasia syndrome, is essential for osteoblast differentiation and bone development. Cell 89:765–771 - PubMed
-
- Enomoto H, Enomoto-Iwamoto M, Iwamoto M, Nomura S, Himeno M, Kitamura Y, Kishimoto T, Komori T 2000 Cbfa1 is a positive regulatory factor in chondrocyte maturation. J Biol Chem 275:8695–8702 - PubMed
-
- Yoshida CA, Komori T 2005 Role of Runx proteins in chondrogenesis. Crit Rev Eukaryot Gene Expr 15:243–254 - PubMed
-
- Ducy P, Zhang R, Geoffroy V, Ridall AL, Karsenty G 1997 Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell 89:747–754 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- DK071122/DK/NIDDK NIH HHS/United States
- R01 AR047052/AR/NIAMS NIH HHS/United States
- C06 CA062528/CA/NCI NIH HHS/United States
- R01 DK071122/DK/NIDDK NIH HHS/United States
- R01 CA109147/CA/NCI NIH HHS/United States
- R56 DK071122/DK/NIDDK NIH HHS/United States
- C06 RR14514-01/RR/NCRR NIH HHS/United States
- C06 RR014514/RR/NCRR NIH HHS/United States
- CA109147/CA/NCI NIH HHS/United States
- AR047052/AR/NIAMS NIH HHS/United States
- P30 CA 014089-30/CA/NCI NIH HHS/United States
- C06 CA62528-01/CA/NCI NIH HHS/United States
- P30 CA014089/CA/NCI NIH HHS/United States
- C06 RR10600-01/RR/NCRR NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
