

Crystallizing membrane proteins using lipidic mesophases
- PMID: 19390528
- PMCID: PMC2732203
- DOI: 10.1038/nprot.2009.31
Crystallizing membrane proteins using lipidic mesophases
Abstract
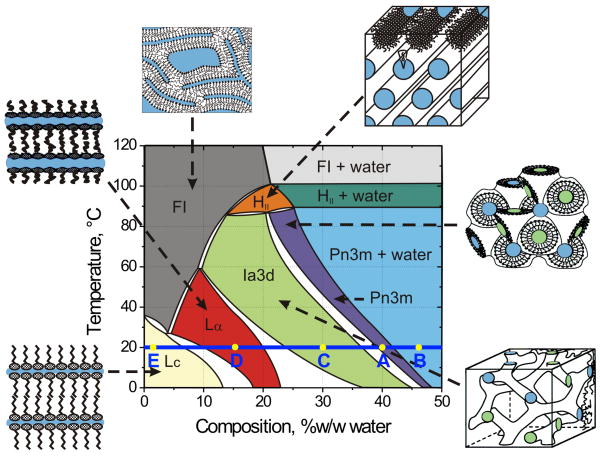
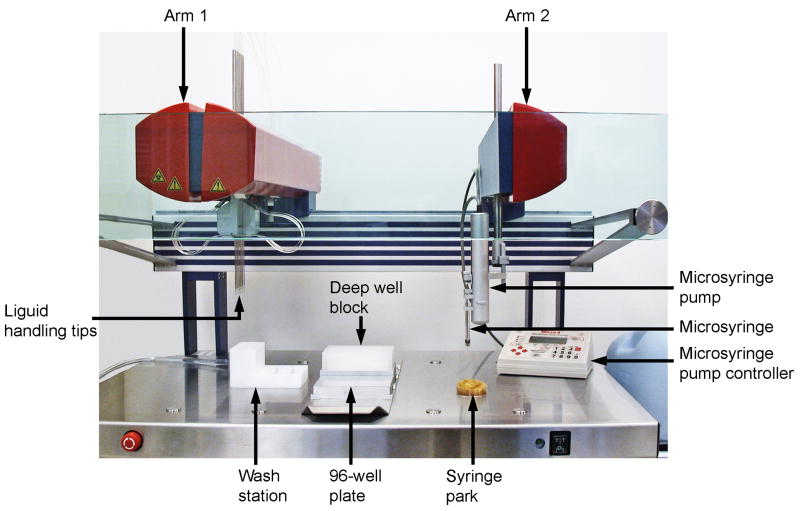
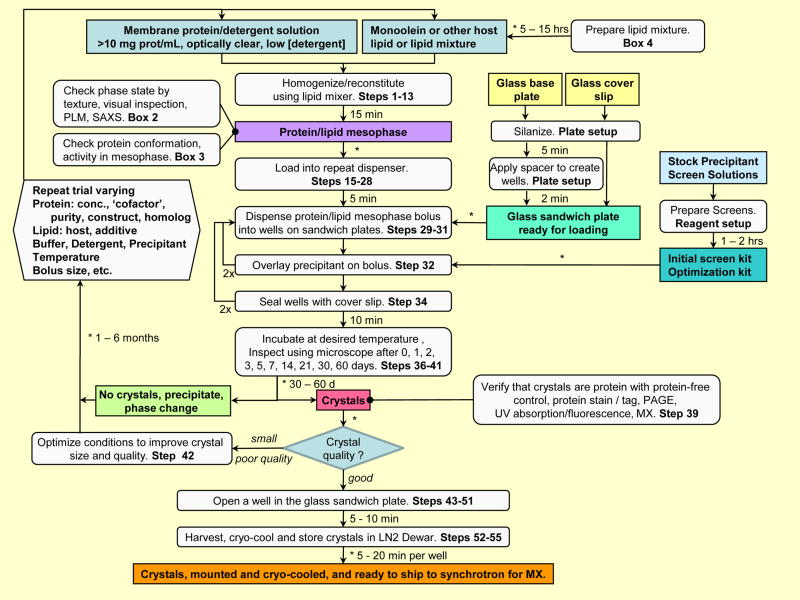
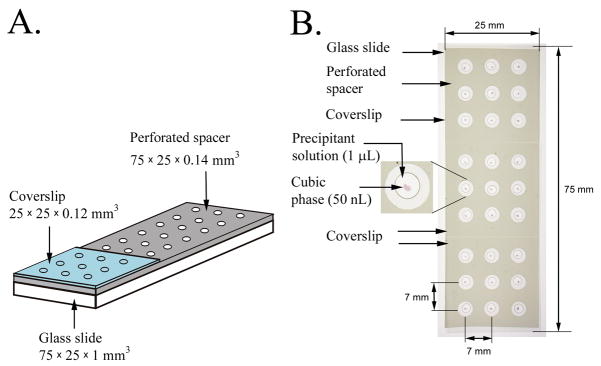
A detailed protocol for crystallizing membrane proteins that makes use of lipidic mesophases is described. This has variously been referred to as the lipid cubic phase or in meso method. The method has been shown to be quite general in that it has been used to solve X-ray crystallographic structures of prokaryotic and eukaryotic proteins, proteins that are monomeric, homo- and hetero-multimeric, chromophore-containing and chromophore-free, and alpha-helical and beta-barrel proteins. Its most recent successes are the human-engineered beta(2)-adrenergic and adenosine A(2A) G protein-coupled receptors. Protocols are provided for preparing and characterizing the lipidic mesophase, for reconstituting the protein into the monoolein-based mesophase, for functional assay of the protein in the mesophase and for setting up crystallizations in manual mode. Methods for harvesting microcrystals are also described. The time required to prepare the protein-loaded mesophase and to set up a crystallization plate manually is about 1 h.
Figures

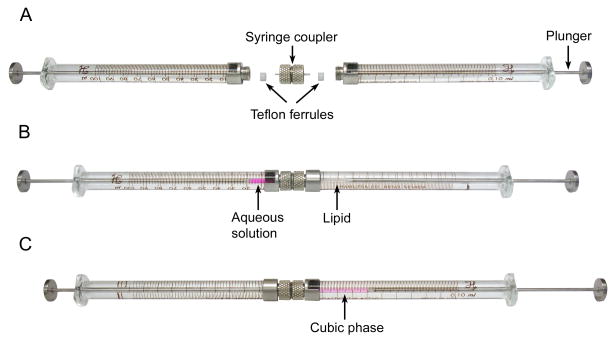
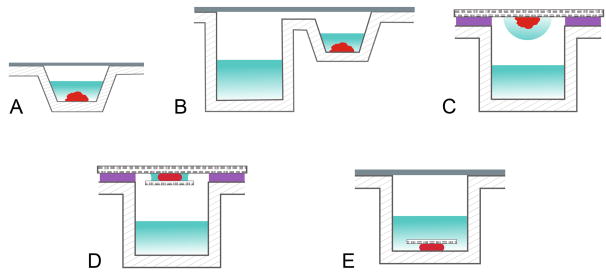
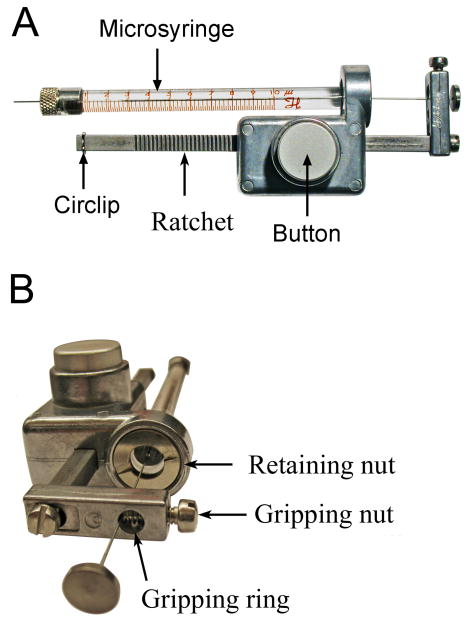
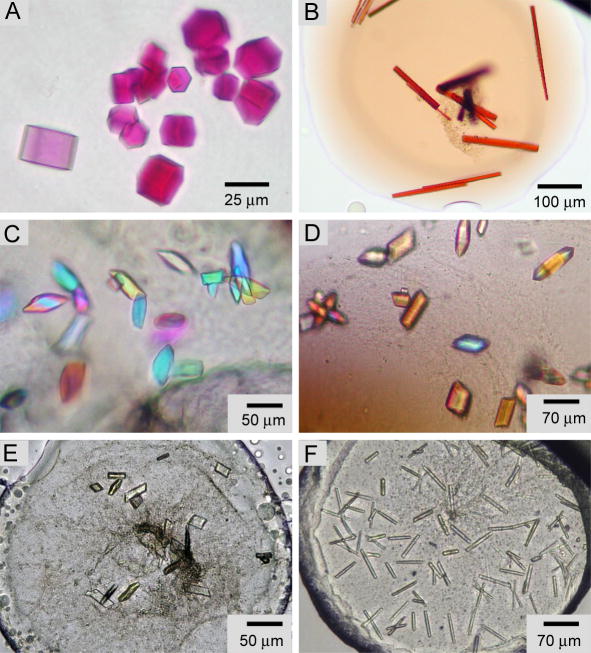
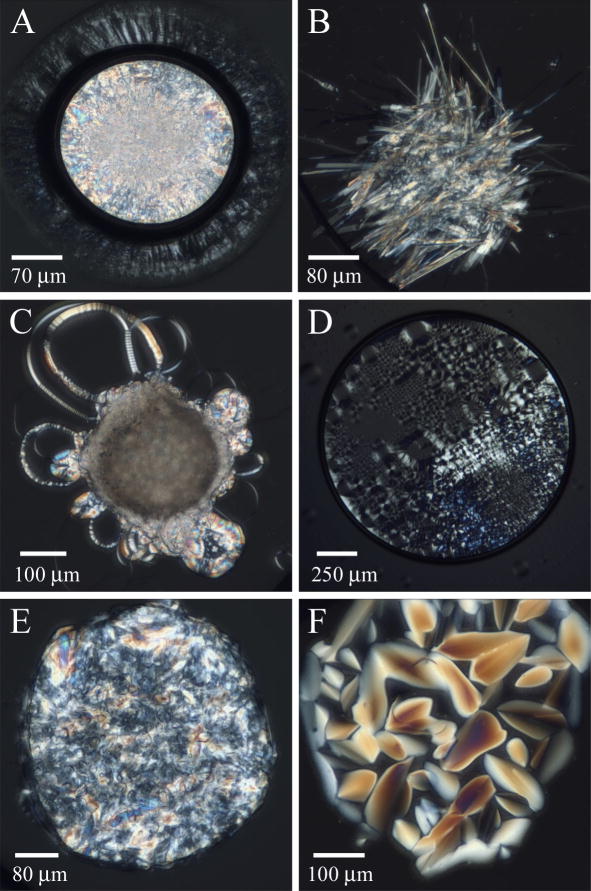
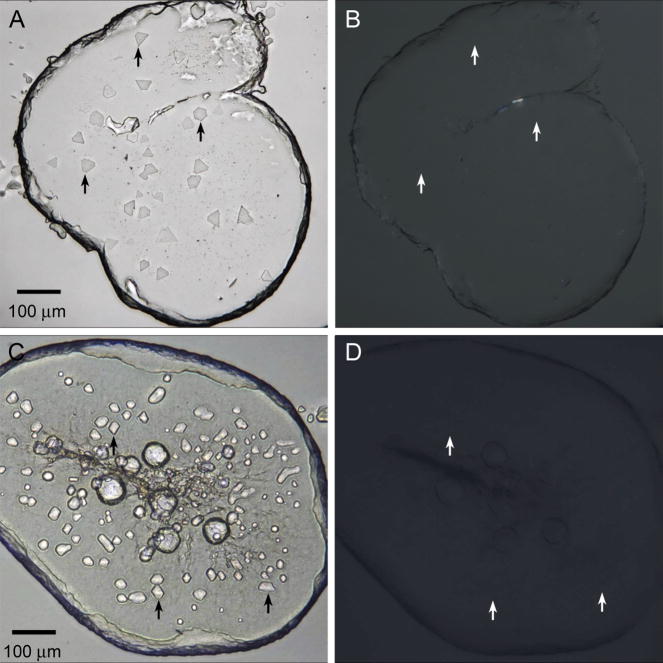
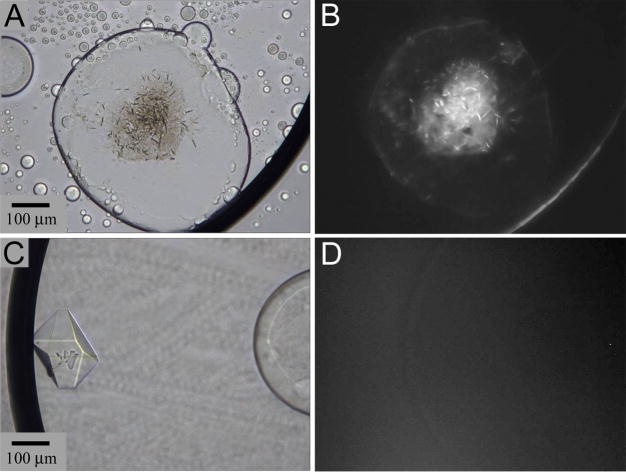

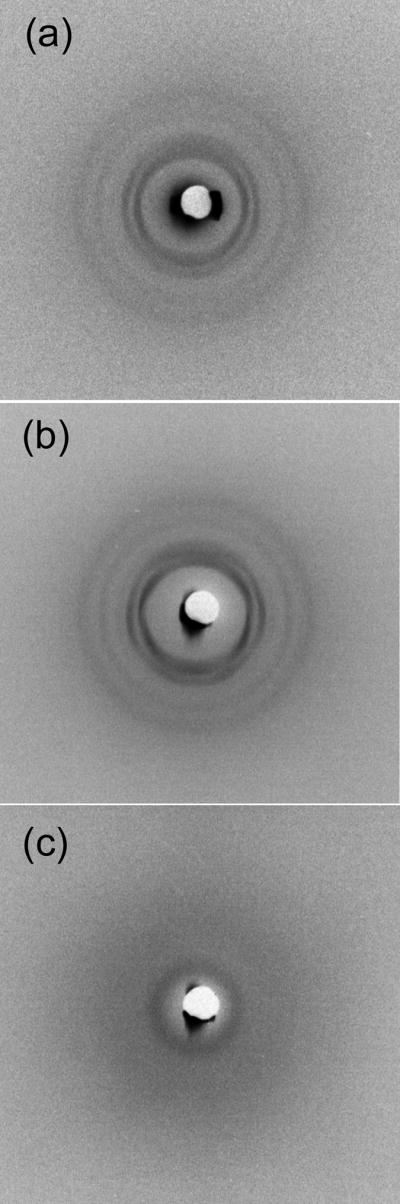
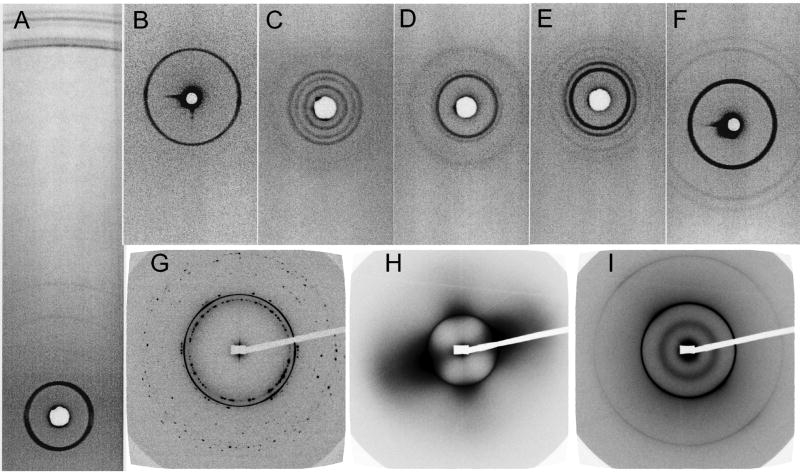

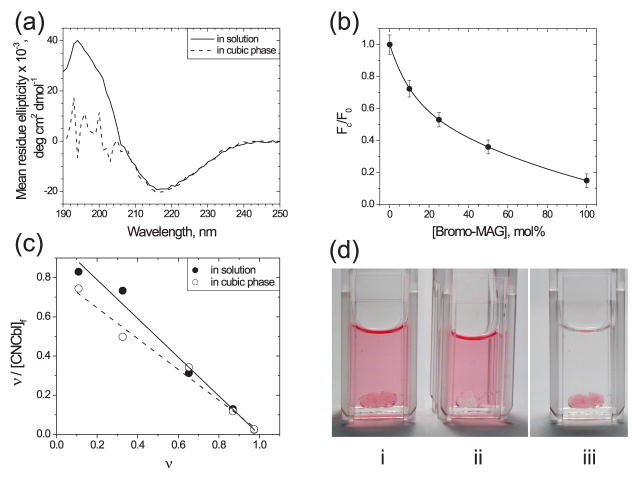
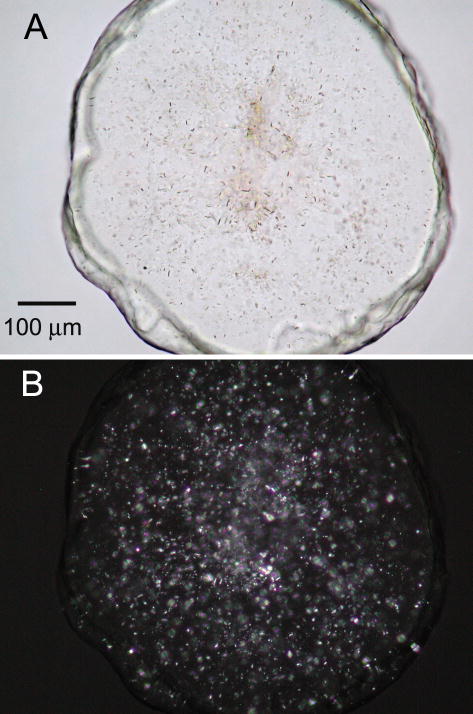
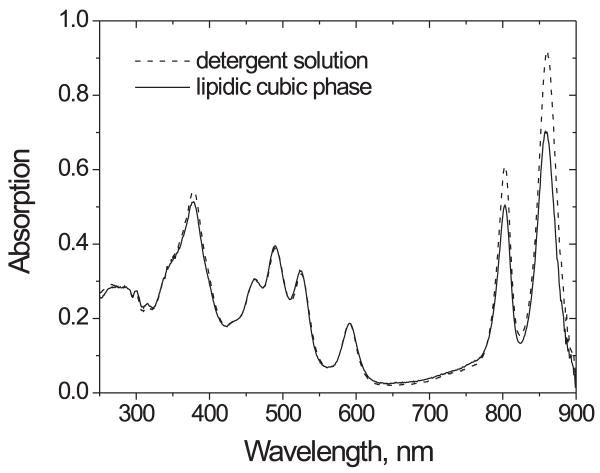
References
-
- Laver WG, Bischofberger N, Webster RG. Disarming flu viruses. Sci Am. 1999;280:78–87. - PubMed
-
- Renfrey S, Featherstone J. Structural proteomics. Nat Rev Drug Discov. 2002;1:175–176. - PubMed
-
- Caffrey M. Membrane protein crystallization. J Struct Biol. 2003;142:108–132. - PubMed
-
- Wiener MC. A pedestrian guide to membrane protein crystallization. Methods. 2004;34:364–372. - PubMed
-
- Sutton BJ, Sohi MK. Crystallization of membrane proteins for X-ray analysis. Methods Mol Biol. 1994;27:1–18. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
