

The structure of a receptor with two associating transmembrane domains on the cell surface: integrin alphaIIbbeta3
- PMID: 19394300
- PMCID: PMC2694939
- DOI: 10.1016/j.molcel.2009.02.022
The structure of a receptor with two associating transmembrane domains on the cell surface: integrin alphaIIbbeta3
Abstract
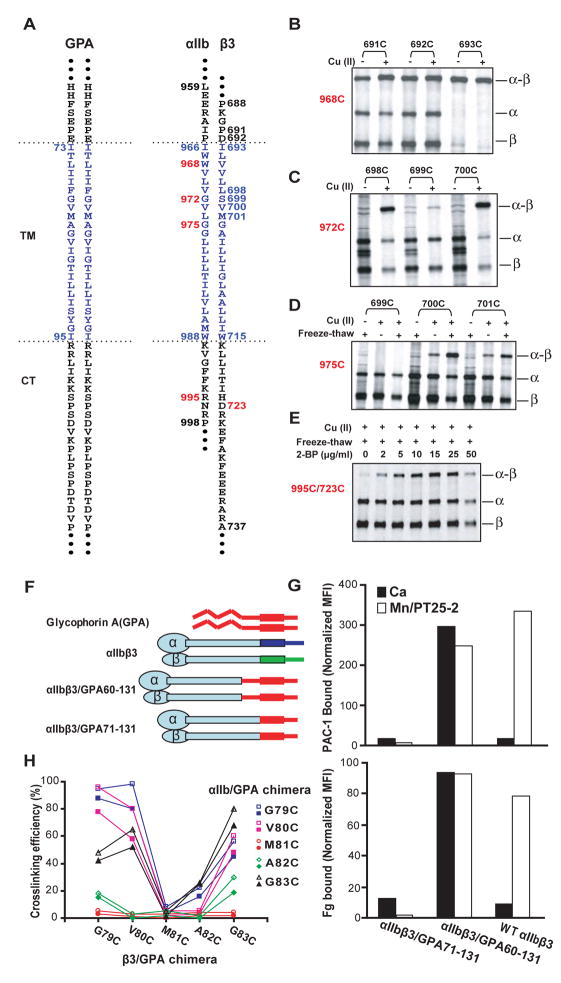
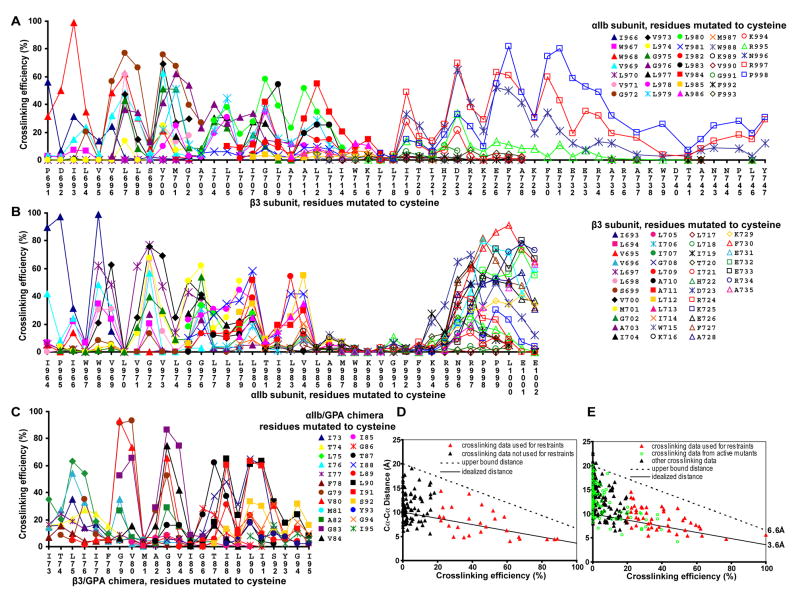
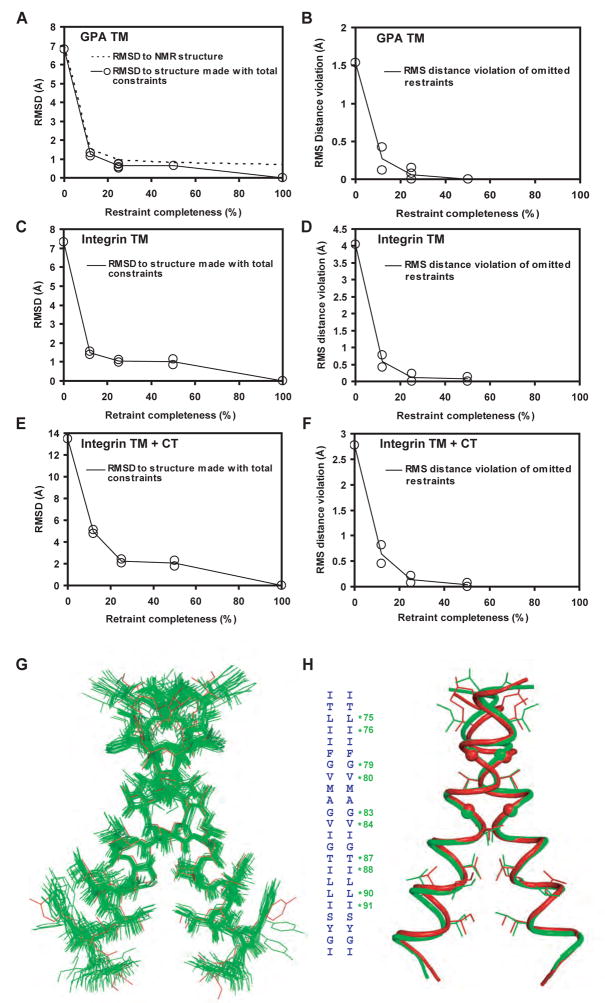
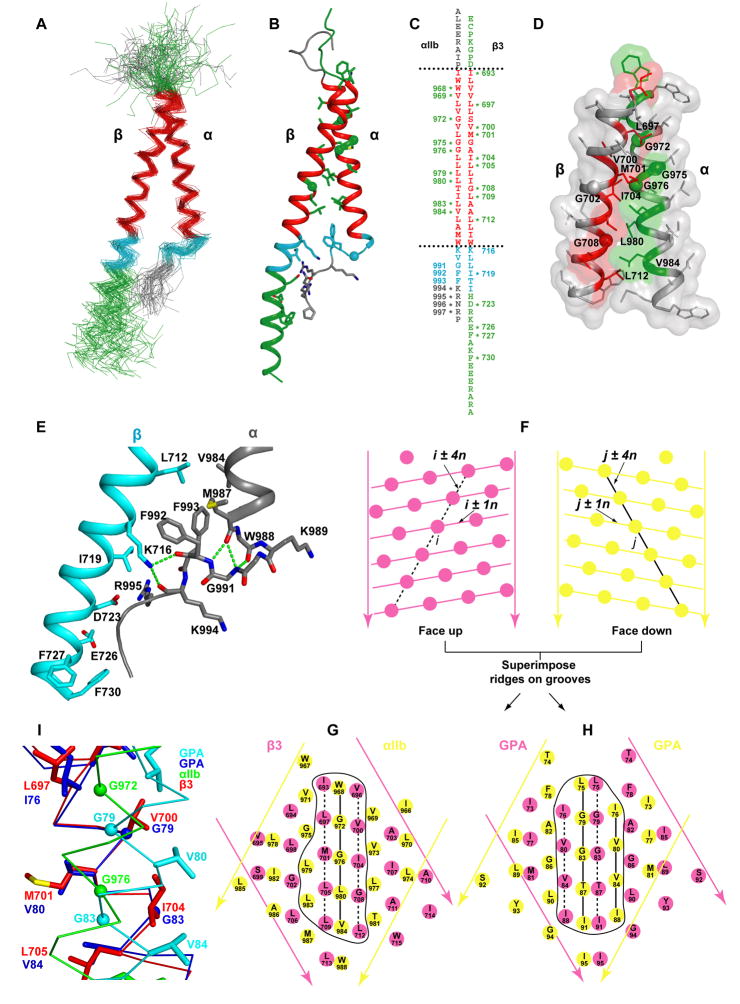
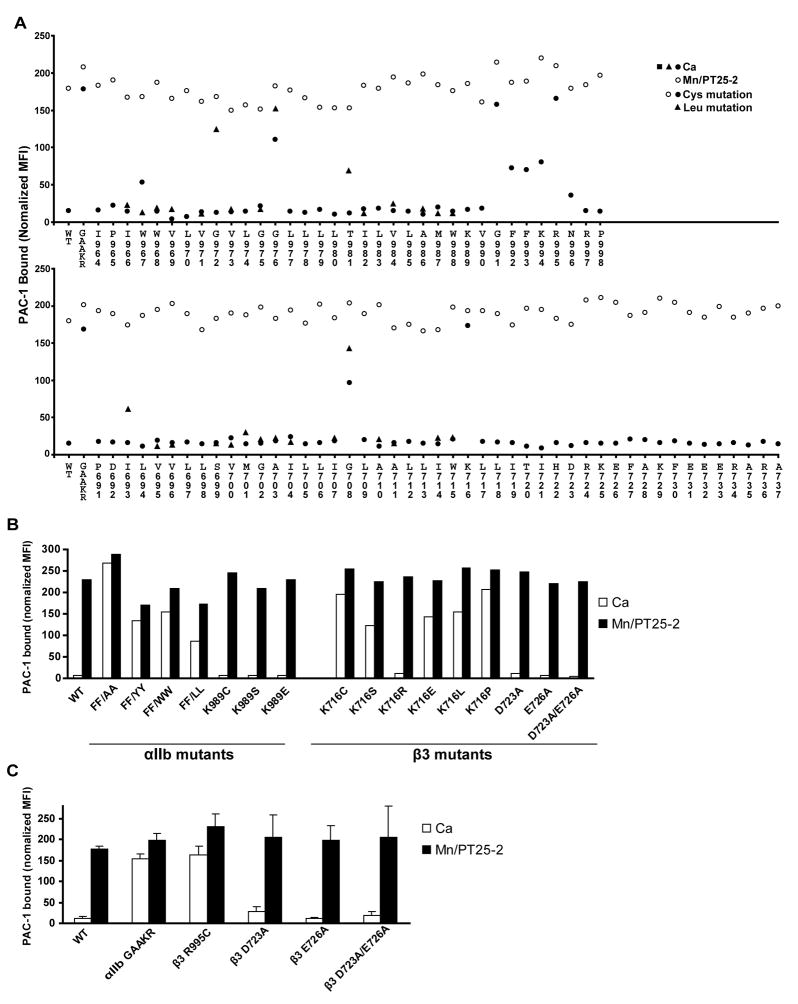
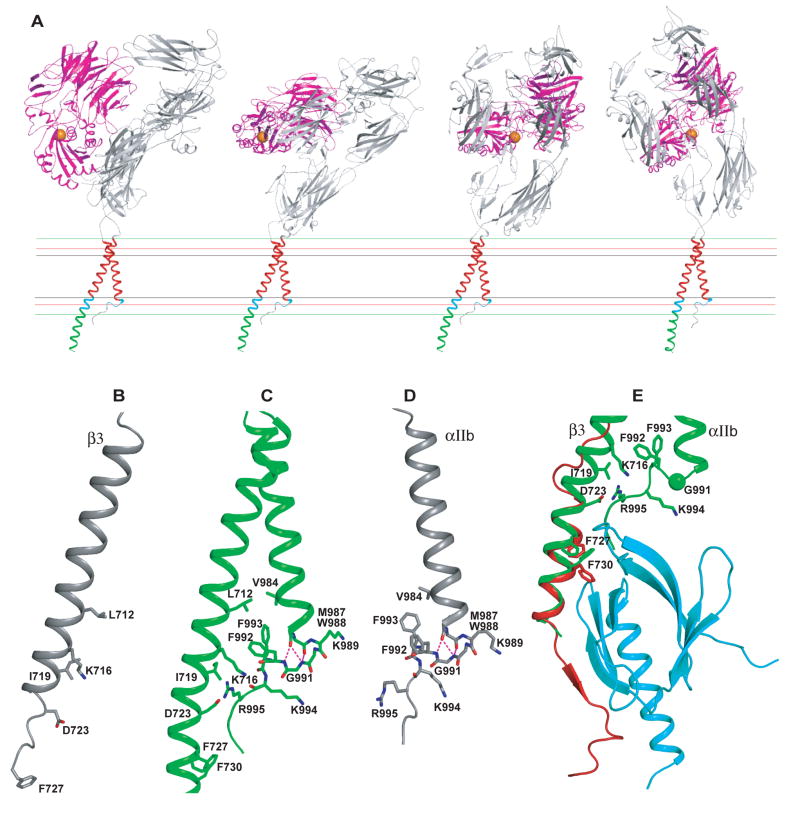
Structures of intact receptors with single-pass transmembrane domains are essential to understand how extracellular and cytoplasmic domains regulate association and signaling through transmembrane domains. A chemical and computational method to determine structures of the membrane regions of such receptors on the cell surface is developed here and validated with glycophorin A. An integrin heterodimer structure reveals association over most of the lengths of the alpha and beta transmembrane domains and shows that the principles governing association of hetero and homo transmembrane dimers differ. A turn at the Gly of the juxtamembrane GFFKR motif caps the alpha TM helix and brings the two Phe of GFFKR into the alpha/beta interface. A juxtamembrane Lys residue in beta also has an important role in the interface. The structure shows how transmembrane association/dissociation regulates integrin signaling. A joint ectodomain and membrane structure shows that substantial flexibility between the extracellular and TM domains is compatible with TM signaling.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
