

A conserved structural motif mediates formation of the periplasmic rings in the type III secretion system
- PMID: 19396170
- PMCID: PMC2860953
- DOI: 10.1038/nsmb.1603
A conserved structural motif mediates formation of the periplasmic rings in the type III secretion system
Abstract
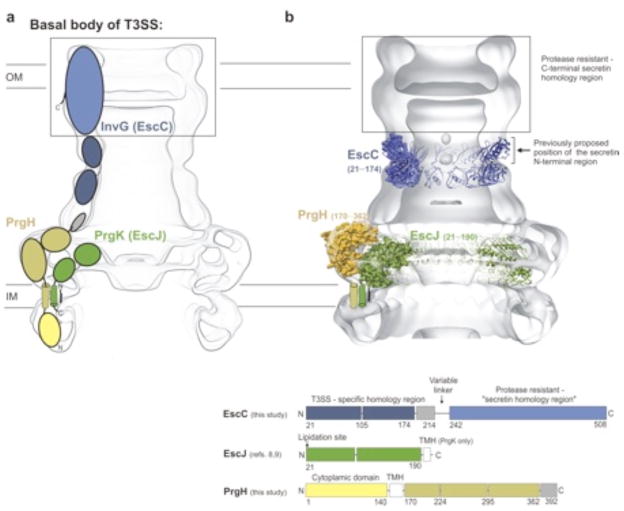
The type III secretion system (T3SS) is a macromolecular 'injectisome' that allows bacterial pathogens to transport virulence proteins into the eukaryotic host cell. This macromolecular complex is composed of connected ring-like structures that span both bacterial membranes. The crystal structures of the periplasmic domain of the outer membrane secretin EscC and the inner membrane protein PrgH reveal the conservation of a modular fold among the three proteins that form the outer membrane and inner membrane rings of the T3SS. This leads to the hypothesis that this conserved fold provides a common ring-building motif that allows for the assembly of the variably sized outer membrane and inner membrane rings characteristic of the T3SS. Using an integrated structural and experimental approach, we generated ring models for the periplasmic domain of EscC and placed them in the context of the assembled T3SS, providing evidence for direct interaction between the outer membrane and inner membrane ring components and an unprecedented span of the outer membrane secretin.
Figures




Comment in
-
Going round in circles: the structural biology of type III secretion systems.Nat Struct Mol Biol. 2009 May;16(5):459-60. doi: 10.1038/nsmb0509-459. Nat Struct Mol Biol. 2009. PMID: 19421157 No abstract available.
References
-
- Kimbrough TG, Miller SI. Assembly of the type III secretion needle complex of Salmonella typhimurium. Microbes Infect. 2002;4:75–82. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
