

Active establishment of centromeric CENP-A chromatin by RSF complex
- PMID: 19398759
- PMCID: PMC2700388
- DOI: 10.1083/jcb.200903088
Active establishment of centromeric CENP-A chromatin by RSF complex
Abstract
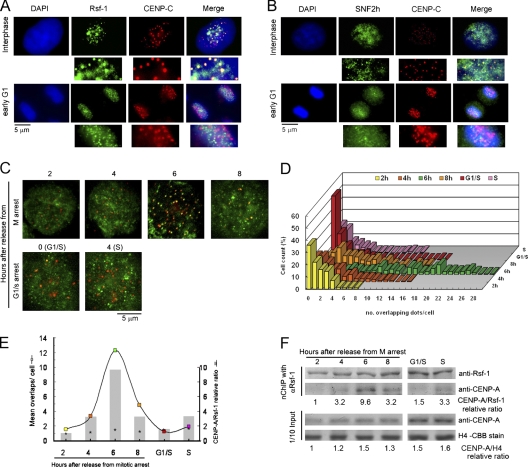
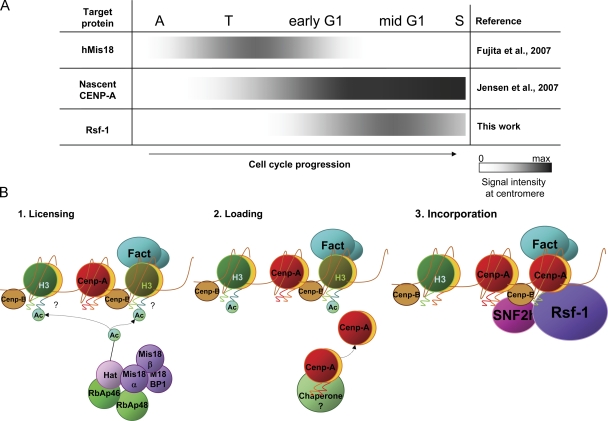
Centromeres are chromosomal structures required for equal DNA segregation to daughter cells, comprising specialized nucleosomes containing centromere protein A (CENP-A) histone, which provide the basis for centromeric chromatin assembly. Discovery of centromere protein components is progressing, but knowledge related to their establishment and maintenance remains limited. Previously, using anti-CENP-A native chromatin immunoprecipitation, we isolated the interphase-centromere complex (ICEN). Among ICEN components, subunits of the remodeling and spacing factor (RSF) complex, Rsf-1 and SNF2h proteins, were found. This paper describes the relationship of the RSF complex to centromere structure and function, demonstrating its requirement for maintenance of CENP-A at the centromeric core chromatin in HeLa cells. The RSF complex interacted with CENP-A chromatin in mid-G1. Rsf-1 depletion induced loss of centromeric CENP-A, and purified RSF complex reconstituted and spaced CENP-A nucleosomes in vitro. From these data, we propose the RSF complex as a new factor actively supporting the assembly of CENP-A chromatin.
Figures




References
-
- Belotserkovskaya R., Oh S., Bonarenko V.A., Orphanides B., Studitsky V.M., Reinberg D. 2003. FACT facilitates transcription-dependent nucleosome alteration.Science. 301:1090–1093 - PubMed
-
- Black B.E., Bassett E.A. 2008. The histone variant CENP-A and centromere specification.Curr. Opin. Cell Biol. 20:91–100 - PubMed
-
- Black B.E., Foltz D.R., Chakravarthy S., Luger K., Woods V.L., Jr., Cleveland D.W. 2004. Structure determinants for generating centromeric chromatin.Nature. 430:578–582 - PubMed
-
- Buchwitz B.J., Ahmad K., Moore L.L., Roth M.B., Henikoff S. 1999. A histone-H3-like protein in C. elegans.Nature. 401:547–548 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
