

Intracellular protein degradation in mammalian cells: recent developments
- PMID: 19399586
- PMCID: PMC11115841
- DOI: 10.1007/s00018-009-0030-6
Intracellular protein degradation in mammalian cells: recent developments
Abstract
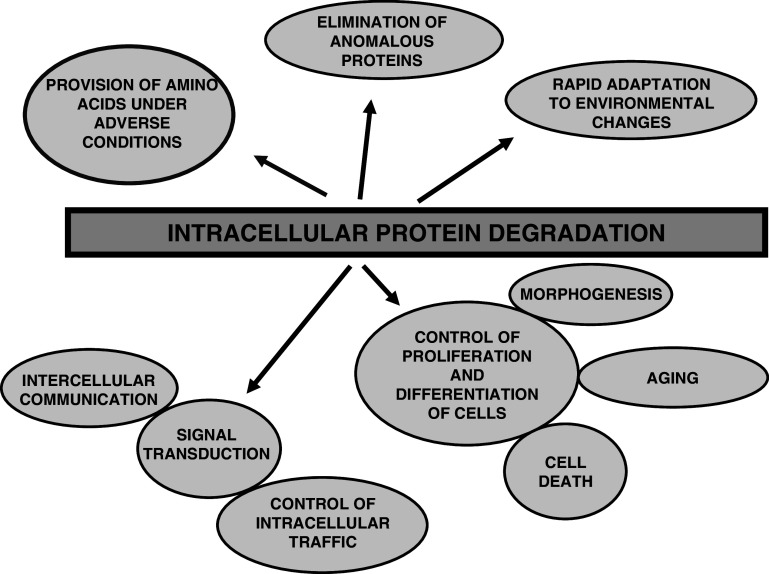
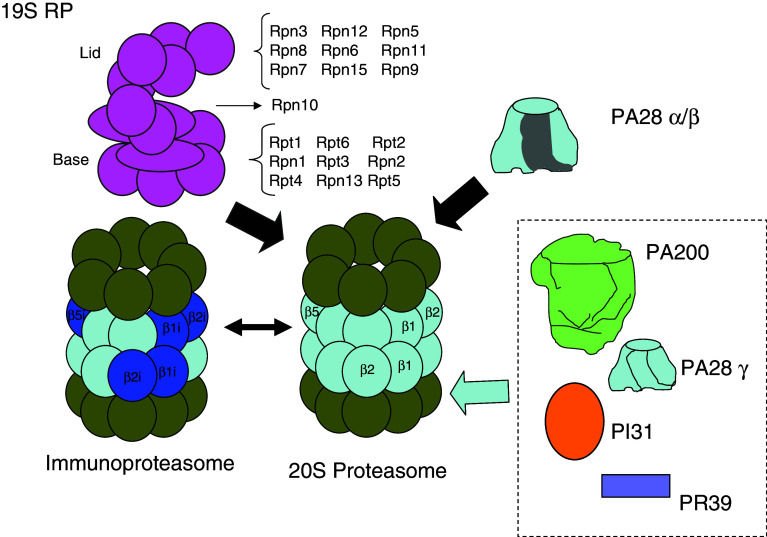
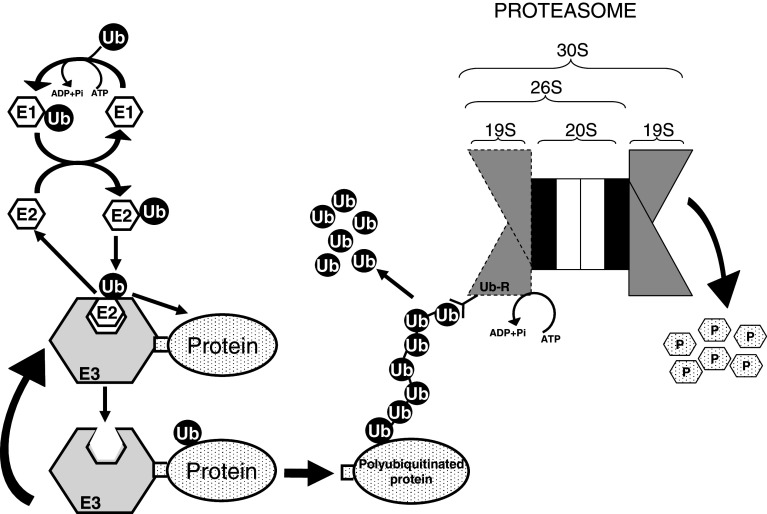
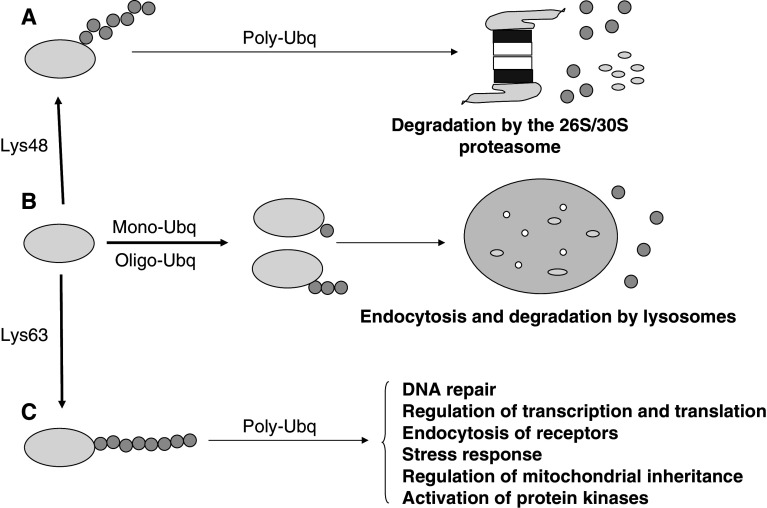
In higher organisms, dietary proteins are broken down into amino acids within the digestive tract but outside the cells, which incorporate the resulting amino acids into their metabolism. However, under certain conditions, an organism loses more nitrogen than is assimilated in the diet. This additional loss was found in the past century to come from intracellular proteins and started an intensive research that produced an enormous expansion of the field and a dispersed literature. Therefore, our purpose is to provide an updated summary of the current knowledge on the proteolytic machinery involved in intracellular protein degradation and its physiological and pathological relevance, especially addressed to newcomers in the field who may find further details in more specialized reviews. However, even providing a general overview, this is an extremely wide field and, therefore, we mainly focus on mammalian cells, while other cells will be mentioned only for comparison purposes.
Figures

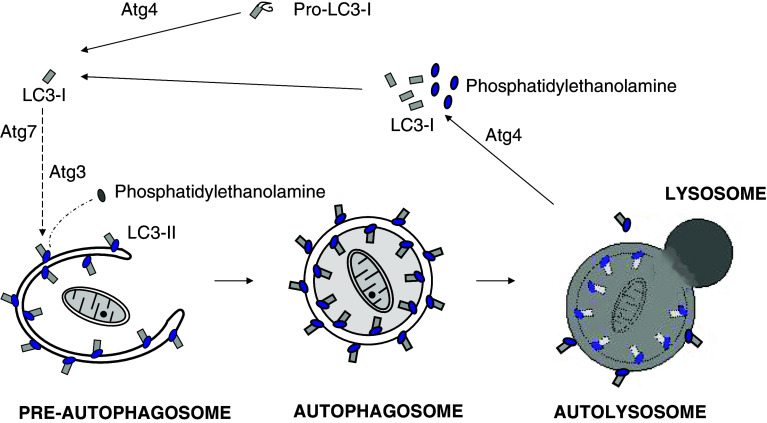
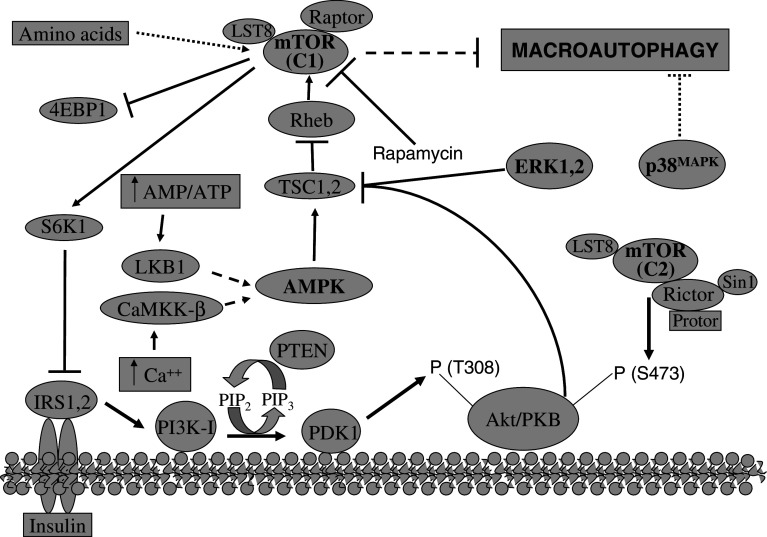
References
-
- Grisolía S, Hernández-Yago J, Knecht E. Regulation of mitochondrial protein concentration: a plausible model which may permit assessing protein turnover. Curr Topics Cell Regul. 1985;27:387–396. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
