

Fluorescent probes for live-cell RNA detection
- PMID: 19400712
- PMCID: PMC2734976
- DOI: 10.1146/annurev-bioeng-061008-124920
Fluorescent probes for live-cell RNA detection
Abstract
Commonly used techniques for analyzing gene expression, such as polymerase chain reaction (PCR), microarrays, and in situ hybridization, have proven invaluable in understanding RNA processing and regulation. However, these techniques rely on the use of lysed and/or fixed cells and are therefore limited in their ability to provide important spatial-temporal information. This has led to the development of numerous techniques for imaging RNA in living cells, some of which have already provided important insight into the dynamic role RNA plays in dictating cell behavior. Here we review the fluorescent probes that have allowed for RNA imaging in living cells and discuss their utility and limitations. Common challenges faced by fluorescent probes, such as probe design, delivery, and target accessibility, are also discussed. It is expected that continued advancements in live cell imaging of RNA will open new and exciting opportunities in a wide range of biological and medical applications.
Figures
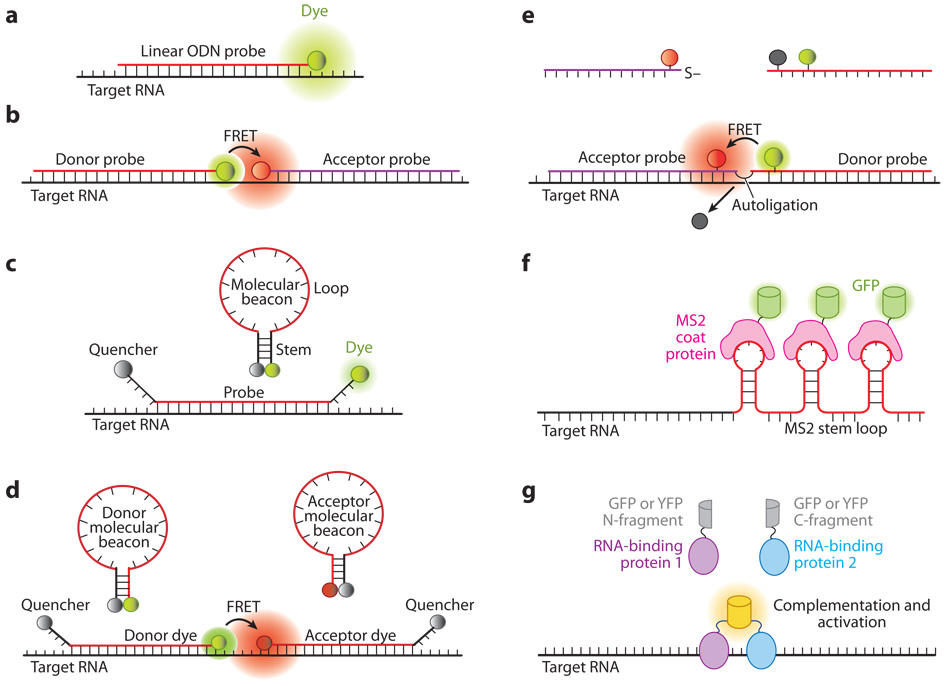

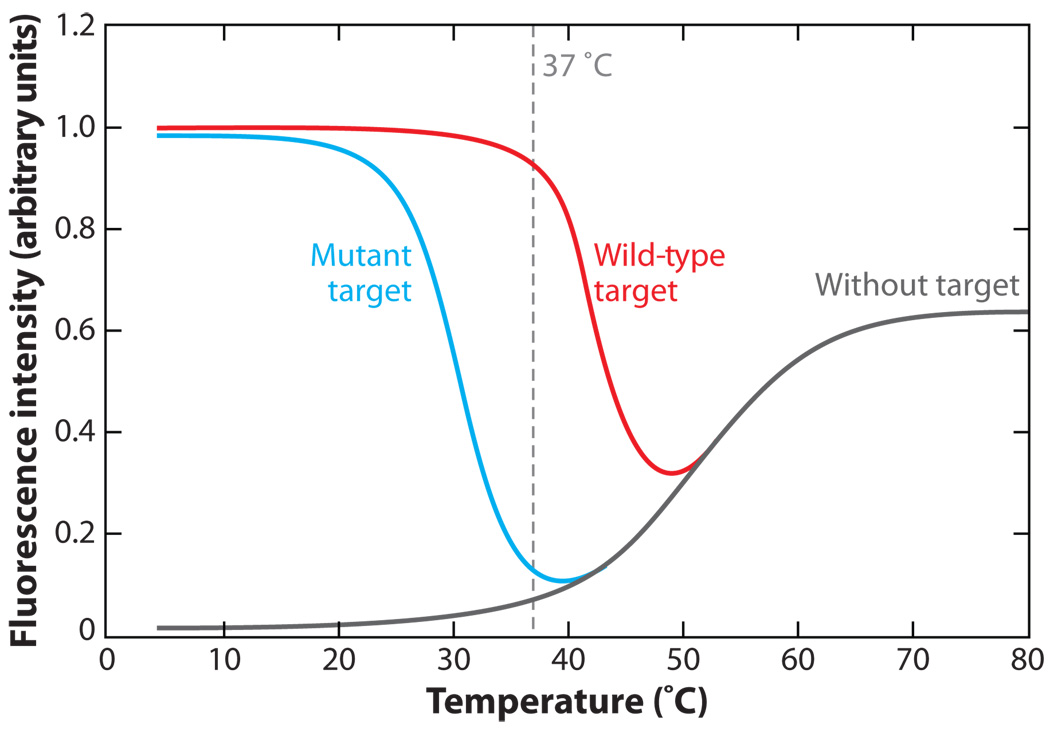
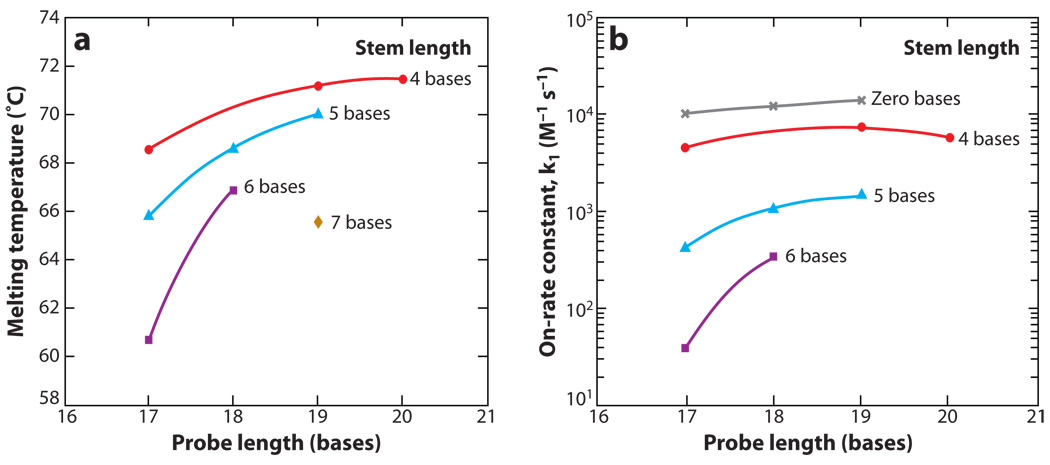
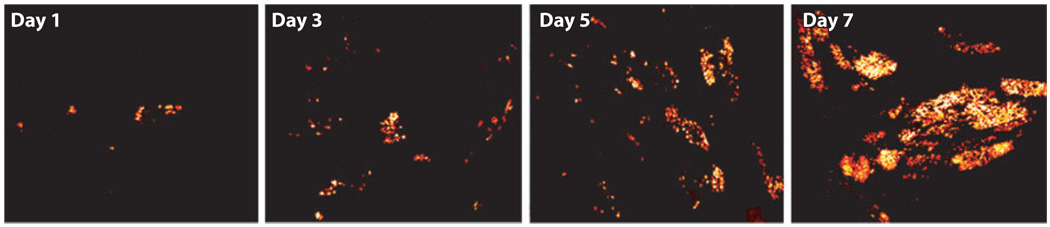
References
-
- Adams MD, Dubnick M, Kerlavage AR, Moreno R, Kelley JM, et al. Sequence identification of 2375 human brain genes. Nature. 1992;355:632–634. - PubMed
-
- Alwine JC, Kemp DJ, Parker BA, Reiser J, Renart J, et al. Detection of specific RNAs or specific fragments of DNA by fractionation in gels and transfer to diazobenzyloxymethyl paper. Methods Enzymol. 1979;68:220–242. - PubMed
-
- Barry MA, Eastman A. Identification of deoxyribonuclease II as an endonuclease involved in apoptosis. Arch. Biochem. Biophys. 1993;300:440–450. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
