

Axon growth and guidance: receptor regulation and signal transduction
- PMID: 19400716
- PMCID: PMC4765433
- DOI: 10.1146/annurev.neuro.051508.135614
Axon growth and guidance: receptor regulation and signal transduction
Abstract
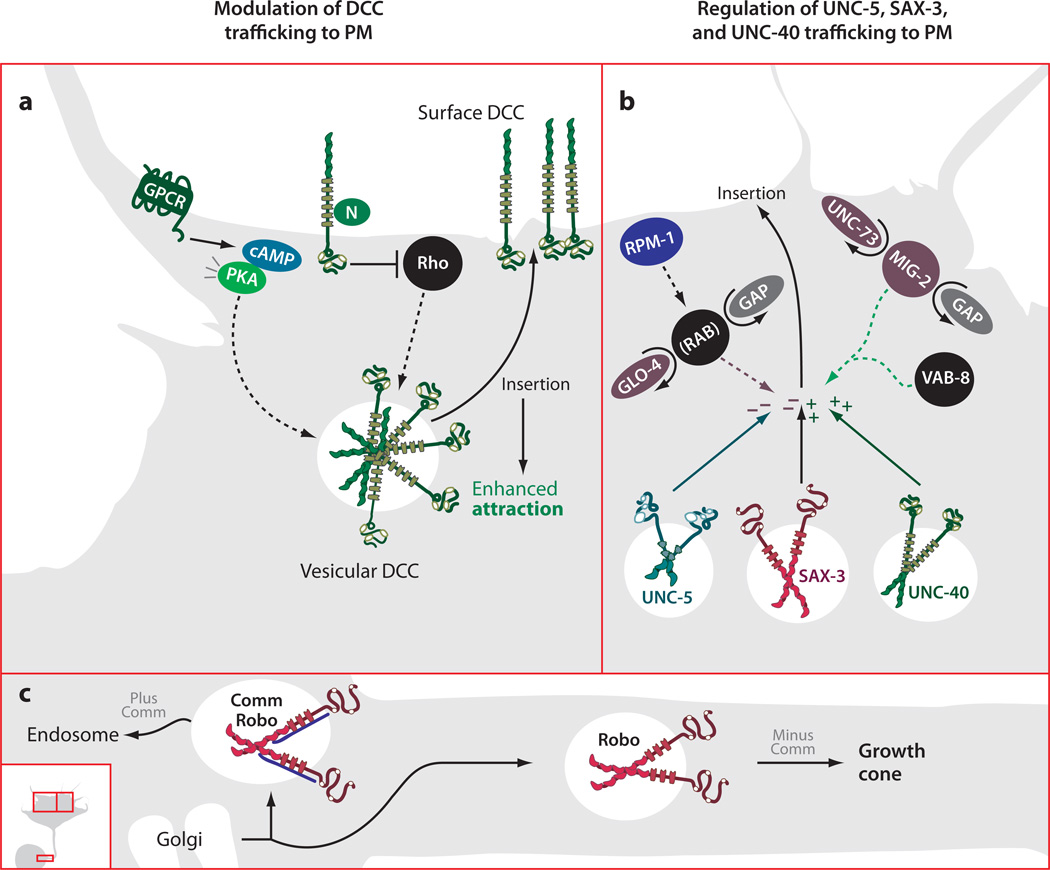
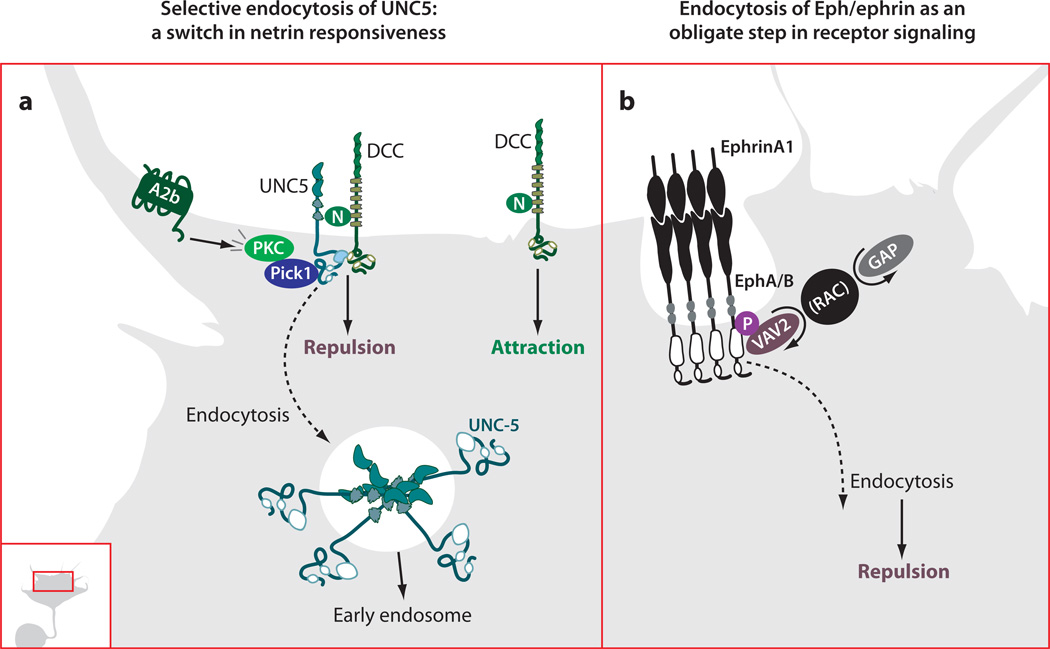
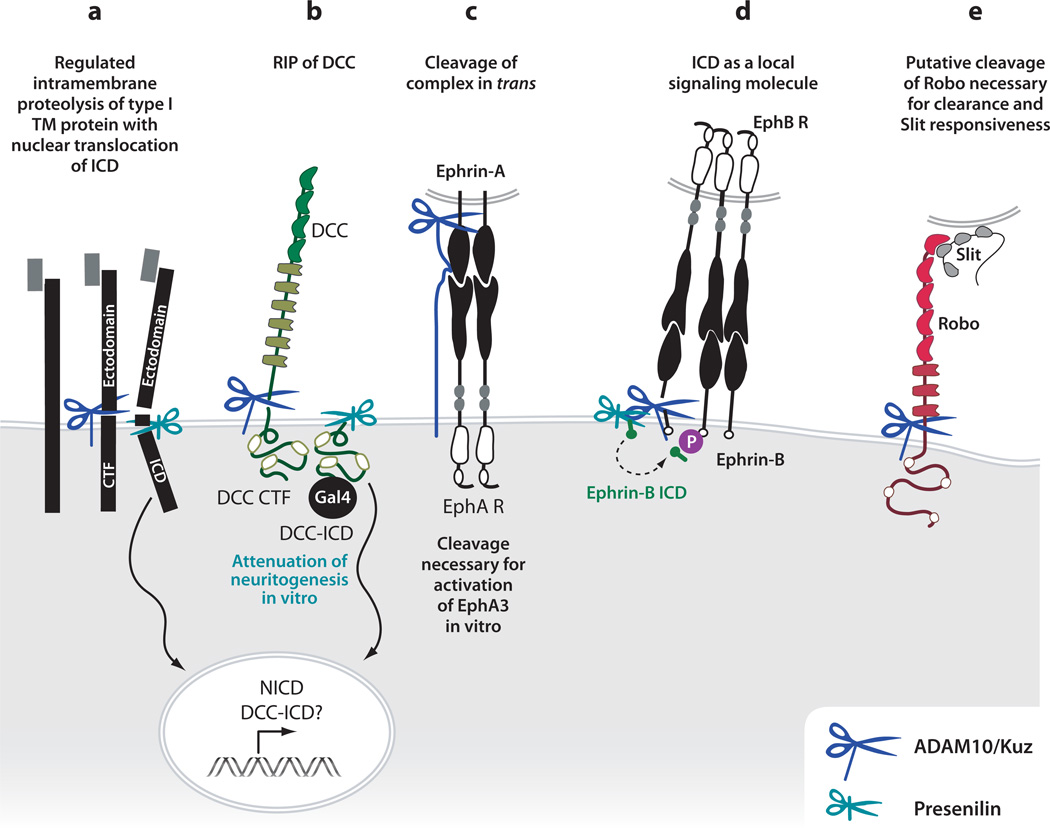
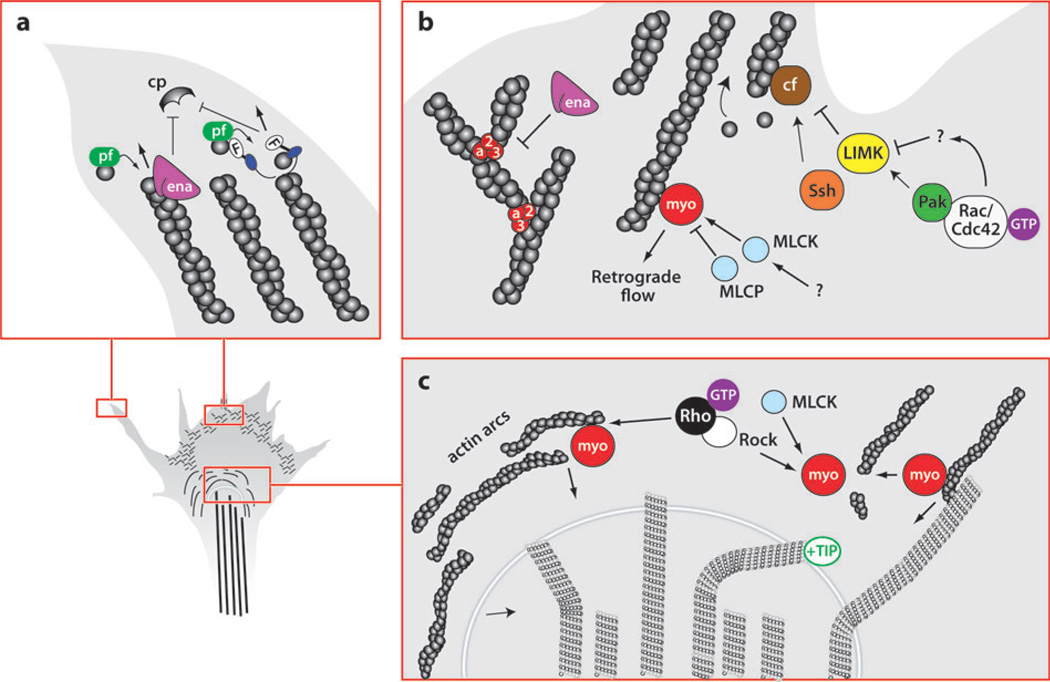
The development of precise connectivity patterns during the establishment of the nervous system depends on the regulated action of diverse, conserved families of guidance cues and their neuronal receptors. Determining how these signaling pathways function to regulate axon growth and guidance is fundamentally important to understanding wiring specificity in the nervous system and will undoubtedly shed light on many neural developmental disorders. Considerable progress has been made in defining the mechanisms that regulate the correct spatial and temporal distribution of guidance receptors and how these receptors in turn signal to the growth cone cytoskeleton to control steering decisions. This review focuses on recent advances in our understanding of the mechanisms mediating growth cone guidance with a particular emphasis on the control of guidance receptor regulation and signaling.
Figures
References
-
- Aizawa H, Wakatsuki S, Ishii A, Moriyama K, Sasaki Y, et al. Phosphorylation of cofilin by LIM-kinase is necessary for semaphorin 3A-induced growth cone collapse. Nat. Neurosci. 2001;4:367–373. - PubMed
-
- Ang LH, Kim J, Stepensky V, Hing H. Dock and Pak regulate olfactory axon pathfinding in Drosophila. Development. 2003;130:1307–1316. - PubMed
-
- Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA, et al. Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature. 1998;393:805–809. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
