

A major cathepsin B protease from the liver fluke Fasciola hepatica has atypical active site features and a potential role in the digestive tract of newly excysted juvenile parasites
- PMID: 19401154
- PMCID: PMC3514016
- DOI: 10.1016/j.biocel.2009.02.003
A major cathepsin B protease from the liver fluke Fasciola hepatica has atypical active site features and a potential role in the digestive tract of newly excysted juvenile parasites
Abstract
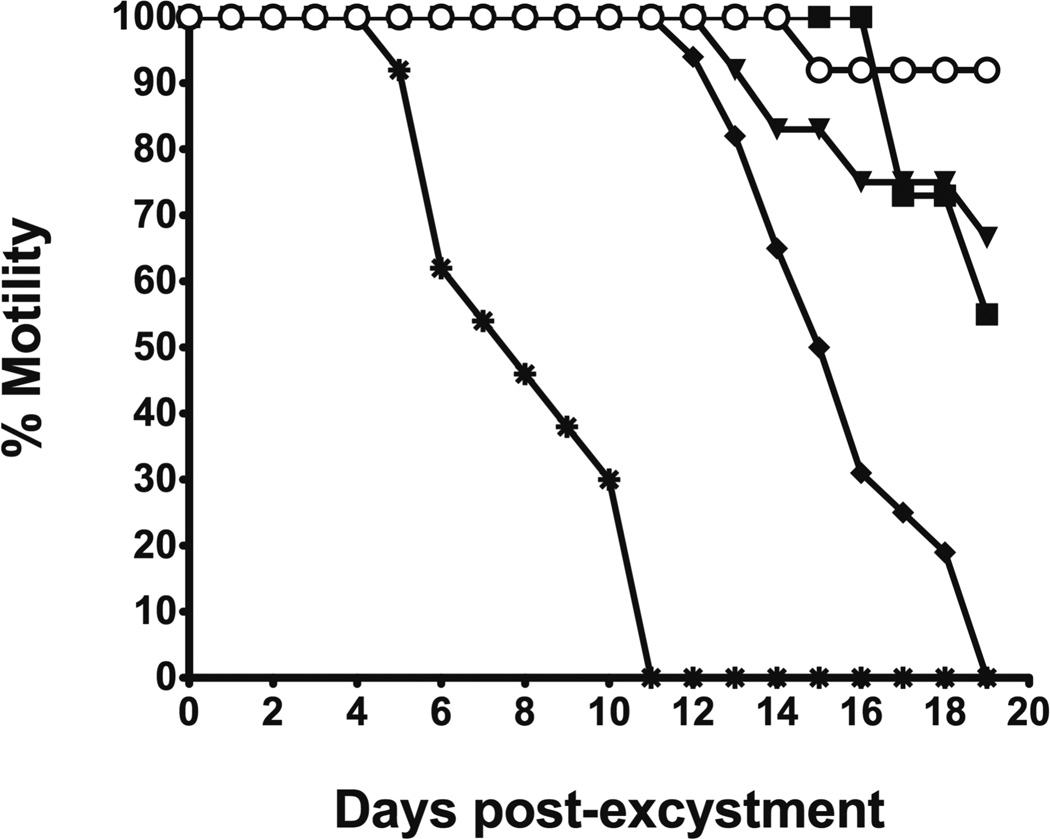
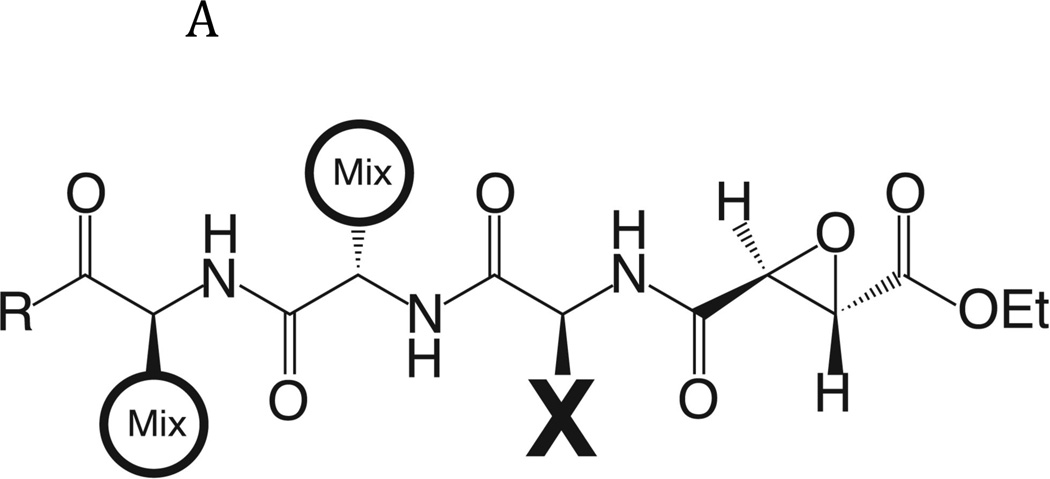
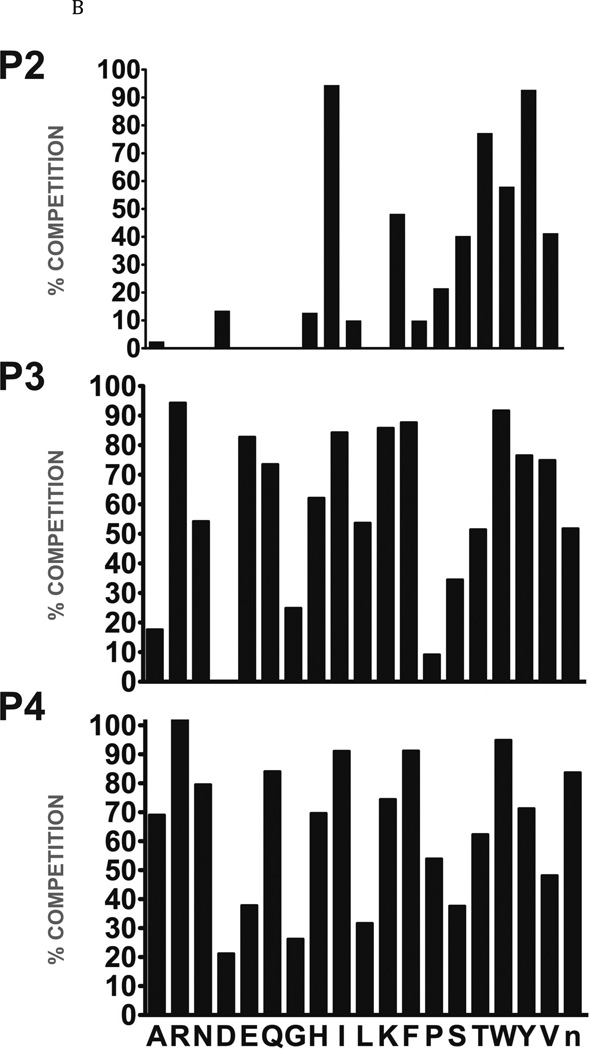
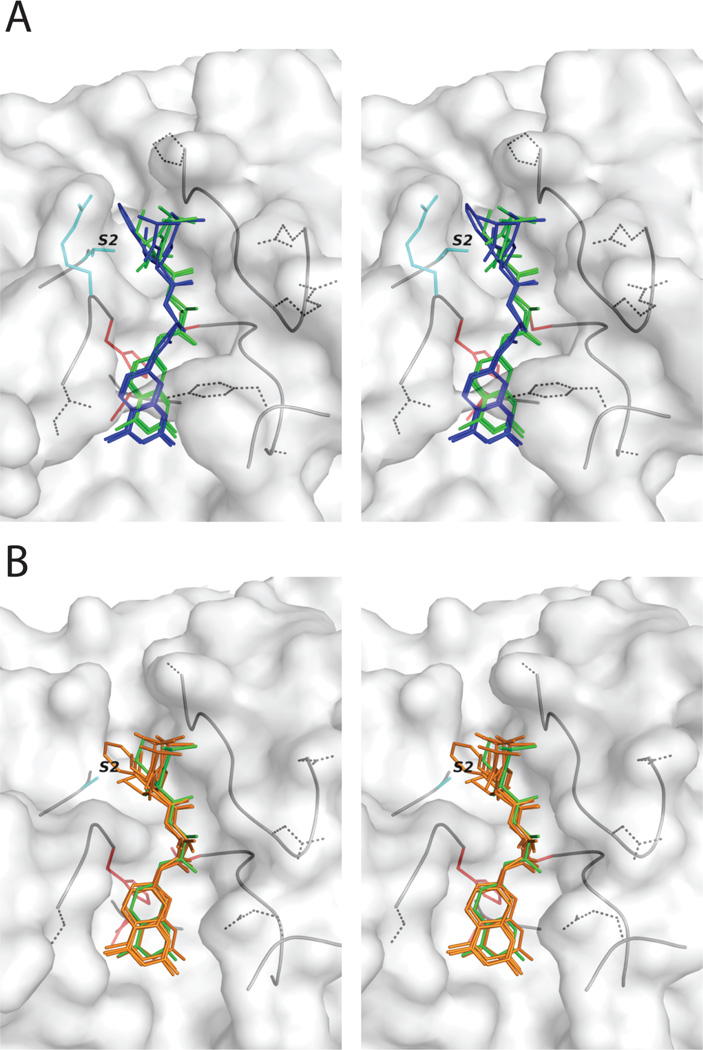
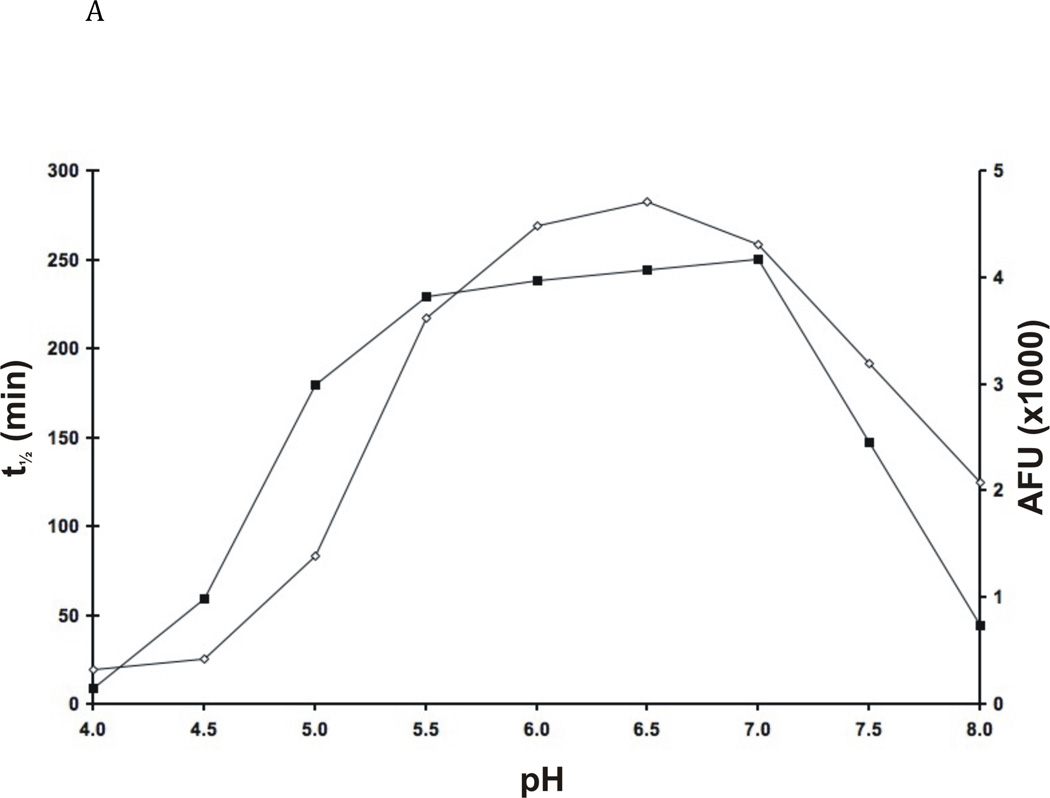
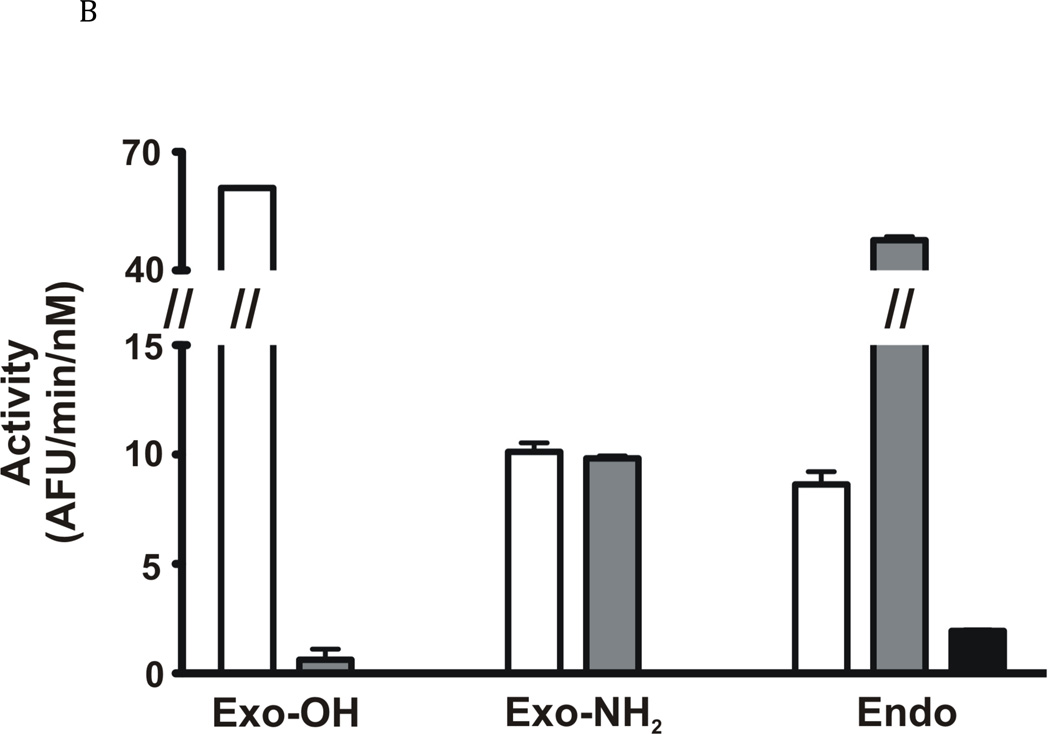
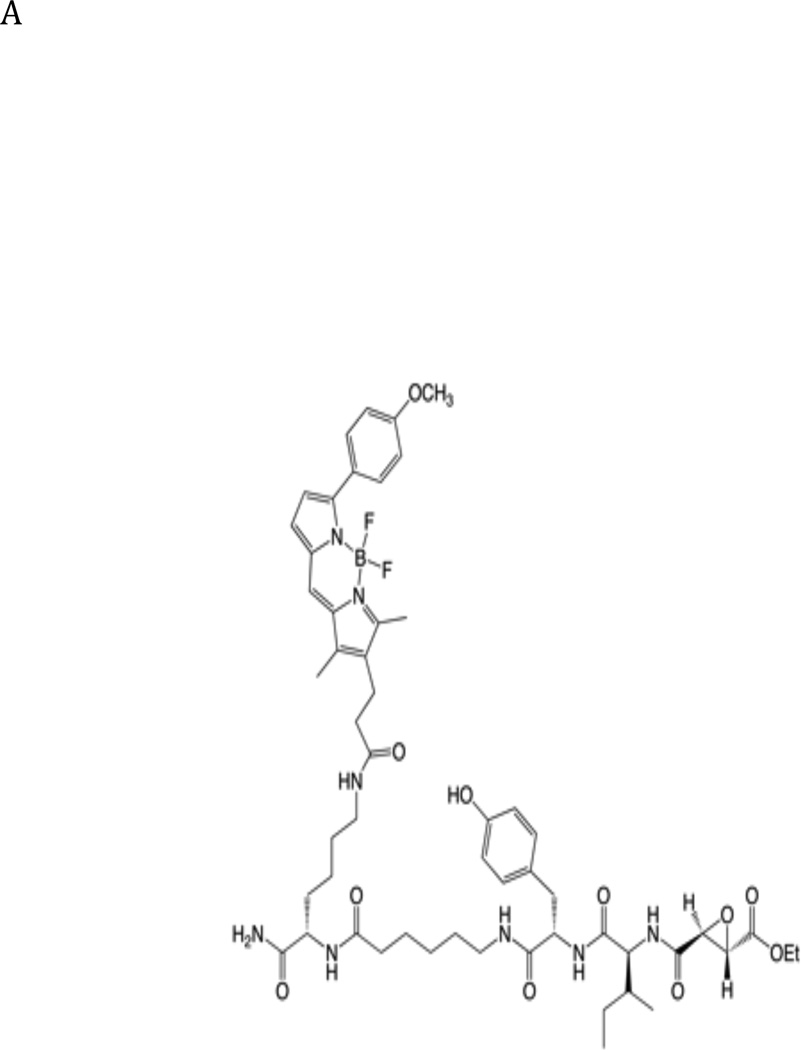
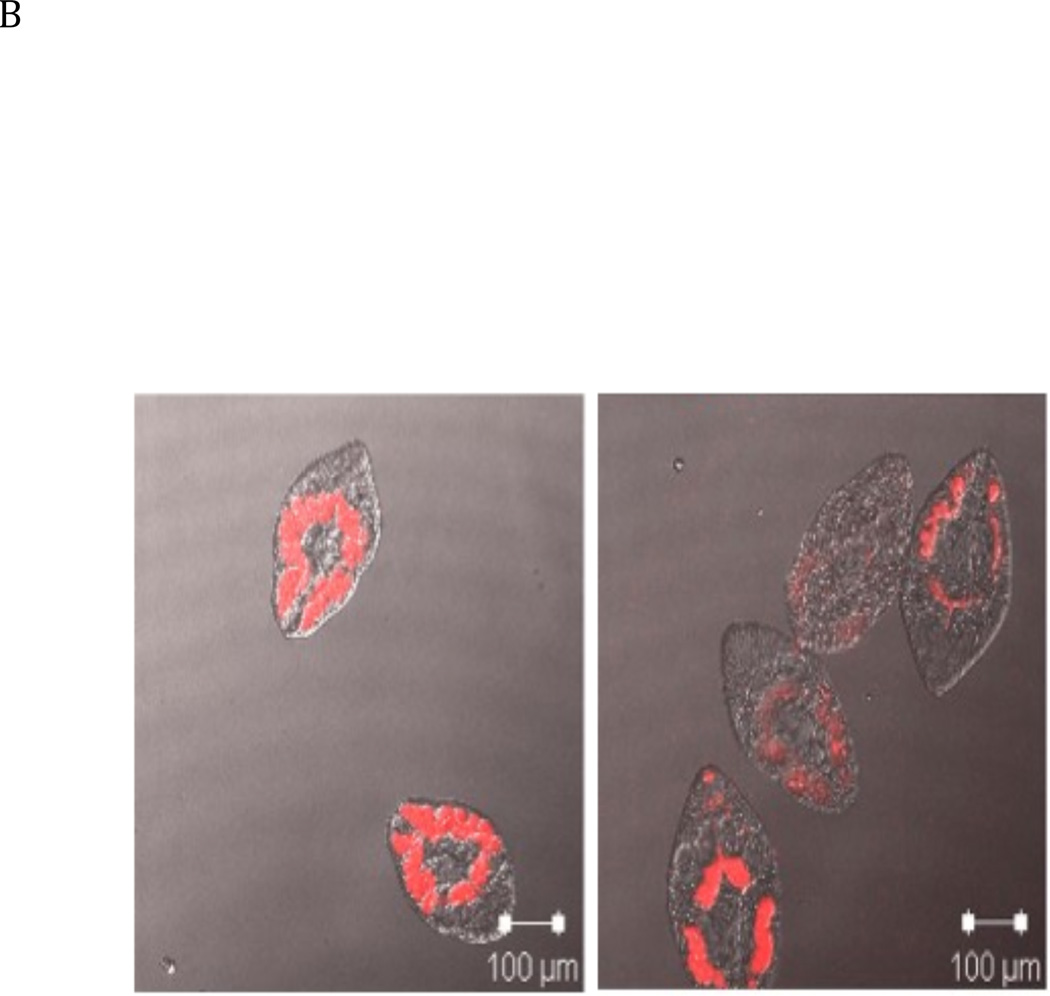
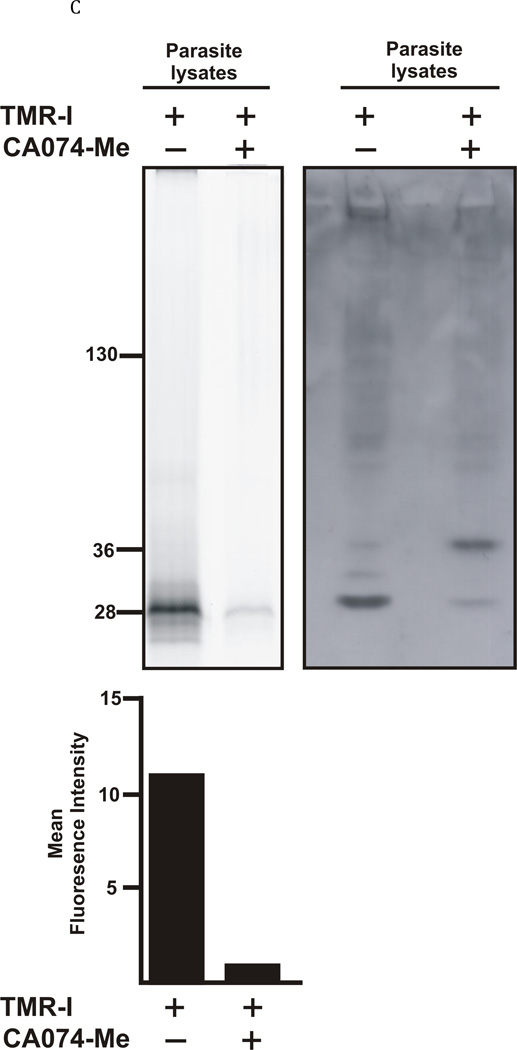
The newly excysted juvenile (NEJ) stage of the Fasciola hepatica lifecycle occurs just prior to invasion into the wall of the gut of the host, rendering it an important target for drug development. The cathepsin B enzymes from NEJ flukes have recently been demonstrated to be crucial to invasion and migration by the parasite. Here we characterize one of the cathepsin B enzymes (recombinant FhcatB1) from NEJ flukes. FhcatB1 has biochemical properties distinct from mammalian cathepsin B enzymes, with an atypical preference for Ile over Leu or Arg residues at the P(2) substrate position and an inability to act as an exopeptidase. FhcatB1 was active across a broad pH range (optimal activity at pH 5.5-7.0) and resistant to inhibition by cystatin family inhibitors from sheep and humans, suggesting that this enzyme would be able to function in extracellular environments in its mammalian hosts. It appears, however, that the FhcatB1 protease functions largely as a digestive enzyme in the gut of the parasite, due to the localization of a specific, fluorescently labeled inhibitor with an Ile at the P(2) position. Molecular modelling and dynamics were used to predict the basis for the unusual substrate specificity: a P(2) Ile residue positions the substrate optimally for interaction with catalytic residues of the enzyme, and the enzyme lacks an occluding loop His residue crucial for exopeptidase activity. The unique features of the enzyme, particularly with regard to its specificity and likely importance to a vital stage of the parasite's life cycle, make it an excellent target for therapeutic inhibitors or vaccination.
Figures

Similar articles
-
Fasciola hepatica: characterization and cloning of the major cathepsin B protease secreted by newly excysted juvenile liver fluke.Exp Parasitol. 1998 Feb;88(2):85-94. doi: 10.1006/expr.1998.4234. Exp Parasitol. 1998. PMID: 9538862
-
A distinctive repertoire of cathepsins is expressed by juvenile invasive Fasciola hepatica.Biochimie. 2008 Oct;90(10):1461-75. doi: 10.1016/j.biochi.2008.04.020. Epub 2008 Jun 5. Biochimie. 2008. PMID: 18573308
-
Production and processing of a recombinant Fasciola hepatica cathepsin B-like enzyme (FhcatB1) reveals potential processing mechanisms in the parasite.Biol Chem. 2006 Aug;387(8):1053-61. doi: 10.1515/BC.2006.130. Biol Chem. 2006. PMID: 16895475
-
Fasciola hepatica cathepsin L-like proteases: biology, function, and potential in the development of first generation liver fluke vaccines.Int J Parasitol. 2003 Sep 30;33(11):1173-81. doi: 10.1016/s0020-7519(03)00171-1. Int J Parasitol. 2003. PMID: 13678633 Review.
-
Controversial aspects of the life cycle of Fasciola hepatica.Exp Parasitol. 2016 Oct;169:81-9. doi: 10.1016/j.exppara.2016.07.010. Epub 2016 Jul 28. Exp Parasitol. 2016. PMID: 27475124 Review.
Cited by
-
The Effects of pH and Temperature on Cysteine Protease (Cathepsin B) Activity in Miracidia and Eggs of Fasciola hepatica.Iran J Parasitol. 2020 Apr-Jun;15(2):233-239. Iran J Parasitol. 2020. PMID: 32595713 Free PMC article.
-
Cathepsin L Inhibitors with Activity against the Liver Fluke Identified From a Focus Library of Quinoxaline 1,4-di-N-Oxide Derivatives.Molecules. 2019 Jun 26;24(13):2348. doi: 10.3390/molecules24132348. Molecules. 2019. PMID: 31247891 Free PMC article.
-
The Zoonotic Helminth Parasite Fasciola hepatica: Virulence-Associated Cathepsin B and Cathepsin L Cysteine Peptidases Secreted by Infective Newly Excysted Juveniles (NEJ).Animals (Basel). 2021 Dec 8;11(12):3495. doi: 10.3390/ani11123495. Animals (Basel). 2021. PMID: 34944270 Free PMC article.
-
Immunolocalization and immunodetection of the excretory/secretory (ES) antigens of Fasciola gigantica.PLoS One. 2017 Oct 3;12(10):e0185870. doi: 10.1371/journal.pone.0185870. eCollection 2017. PLoS One. 2017. PMID: 28973017 Free PMC article.
-
Drug resistance in liver flukes.Int J Parasitol Drugs Drug Resist. 2020 Apr;12:39-59. doi: 10.1016/j.ijpddr.2019.11.003. Epub 2020 Jan 10. Int J Parasitol Drugs Drug Resist. 2020. PMID: 32179499 Free PMC article. Review.
References
-
- Anon . World Health Organization Technical Series No. 849. Geneva, Switzerland: 1995. Control of Foodborne Trematode Infections. - PubMed
-
- Aronja R, Riancho JA, Aguado JM. Fascioliasis in developed countries: A review of classic and aberrant forms of the disease. Medicine. 1995;74:13–23. - PubMed
-
- Beckham SA, Law RHP, Smooker PM, Quinsey NS, Caffrey CR, McKerrow JH, et al. Production and processing of a recombinant Fasciola hepatica cathepsin B-like enzyme (FhcatB1) reveals potential processing mechanisms in the parasite. Biol Chem. 2006;387:1053–1061. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
