

Lethal pulmonary infection with Francisella novicida is associated with severe sepsis
- PMID: 19401387
- PMCID: PMC2735285
- DOI: 10.1189/jlb.1208728
Lethal pulmonary infection with Francisella novicida is associated with severe sepsis
Abstract
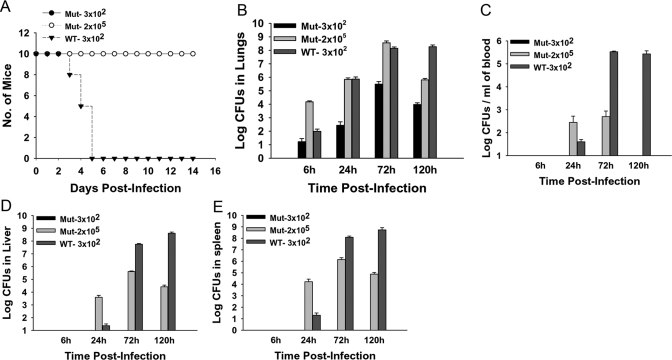
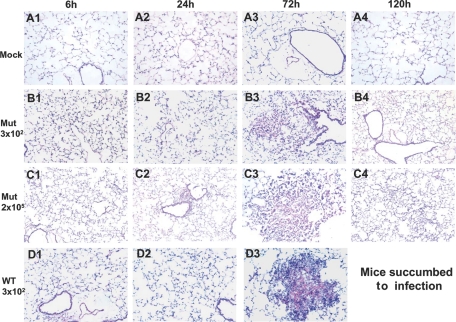
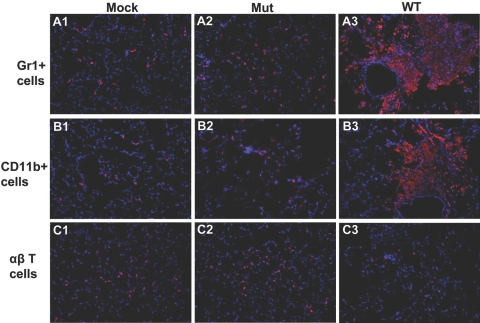
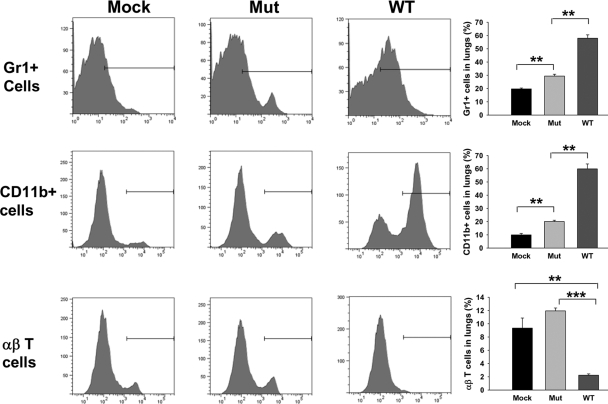
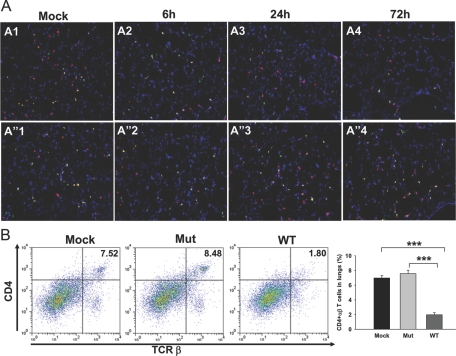
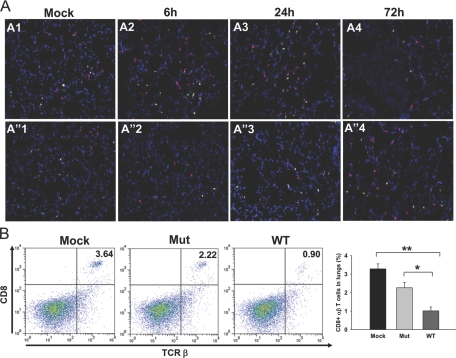
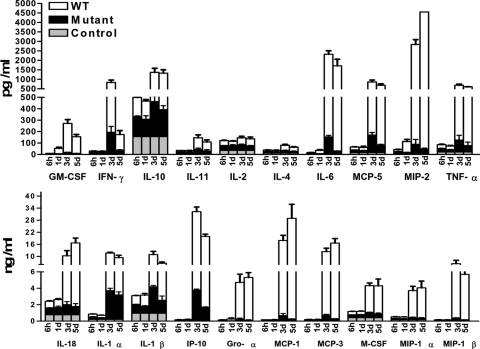
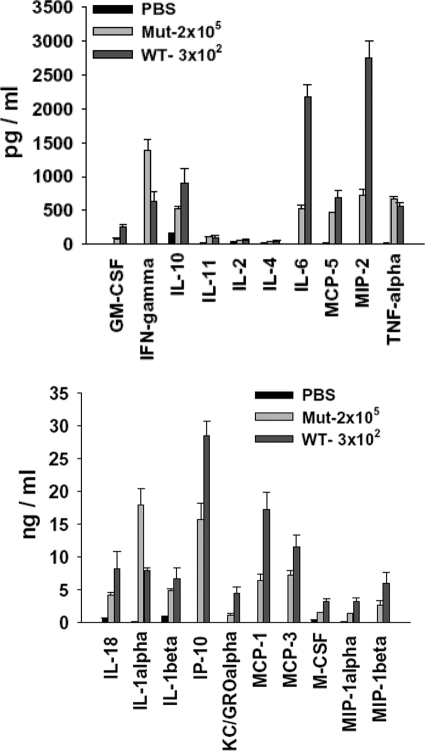
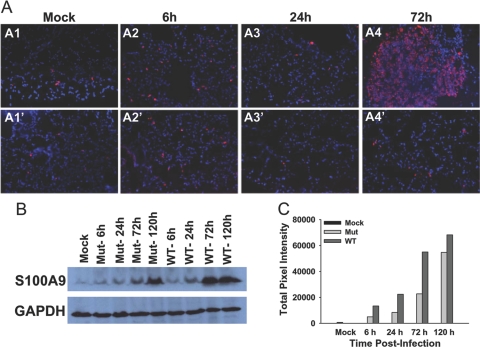
The bacterial or host determinants of lethality associated with respiratory Francisella infections are currently unknown. No exo- or endotoxins that contribute to the severity of this disease have been identified. However, a deregulated host immune response upon infection is characterized by an initial 36- to 48-h delay followed by a rapid and excessive inflammatory response prior to death at 72-120 h. Here, we extend these findings by comparing host immune responses between sublethal and lethal respiratory infections of mice with an attenuated transposon mutant (Mut) of F. novicida (F.n.) strain U112 (sublethal) versus the wild-type (WT) strain (lethal). Infection with WT bacteria, but not the Mut, was characterized by sustained bacteremia and systemic dissemination of the pathogen with temporal increases in bacterial burdens in liver and spleen. Severe pathology with large foci of infiltrates associated with extensive tissue damage was evident in WT-infected lungs, and Mut-infected mice displayed much reduced pathology with intact lung architecture. Similar to other experimental models of severe sepsis, WT- but not the Mut-infected mice exhibited a robust increase in numbers of Gr1+ and CD11b+ cells, while displaying a significant depletion of alphabeta T cells. Further, a dramatic up-regulation of multiple cytokines and chemokines was observed only in lethal WT infection. In addition, an earlier and larger increased expression of S100A9, a known mediator of sepsis, was observed in WT-infected mice. Taken together, these results show that a hyperinflammatory host immune response, culminating in severe sepsis, is responsible for the lethal outcome of respiratory tularemia.
Figures
Comment in
-
Editorial: Proinflammatory cytokines in pneumonic tularemia: too much too late?J Leukoc Biol. 2009 Sep;86(3):469-70. doi: 10.1189/jlb.0309119. J Leukoc Biol. 2009. PMID: 19720615 No abstract available.
References
-
- Dennis D T, Inglesby T V, Henderson D A, Bartlett J G, Ascher M S, Eitzen E, Fine A D, Friedlander A M, Hauer J, Layton M, Lillibridge S R, McDade J E, Osterholm M T, O'Toole T, Parker G, Perl T M, Russell P K, Tonat K, Working Group on Civilian Biodefense Tularemia as a biological weapon: medical and public health management. JAMA. 2001;285:2763–2773. - PubMed
-
- Sjostedt A. Tularemia: history, epidemiology, pathogen physiology, and clinical manifestations. Ann N Y Acad Sci. 2007;1105:1–29. - PubMed
-
- Mares C A, Ojeda S S, Morris E G, Li Q, Teale J M. Initial delay in the immune response to Francisella tularensis is followed by hypercytokinemia characteristic of severe sepsis and correlating with upregulation and release of damage-associated molecular patterns. Infect Immun. 2008;76:3001–3010. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
