

Dynamically driven ligand selectivity in cyclic nucleotide binding domains
- PMID: 19403523
- PMCID: PMC2749143
- DOI: 10.1074/jbc.M109.011700
Dynamically driven ligand selectivity in cyclic nucleotide binding domains
Abstract
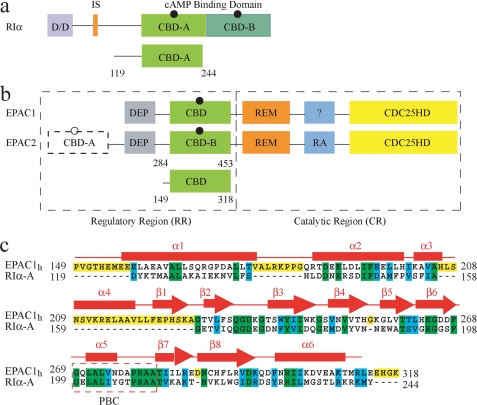
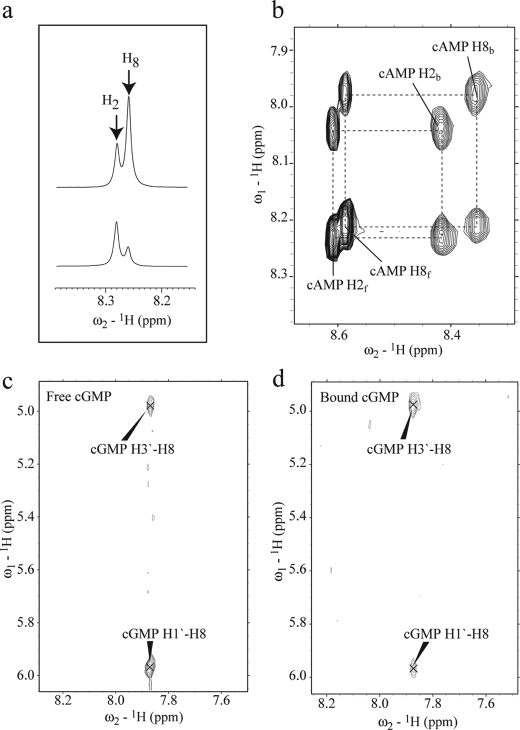
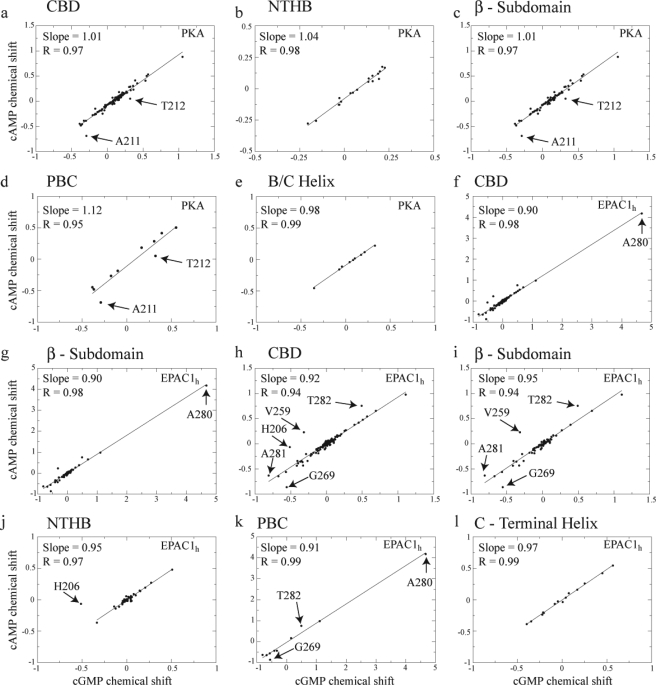
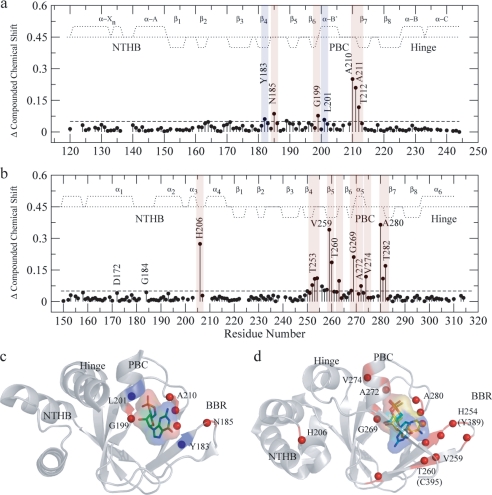
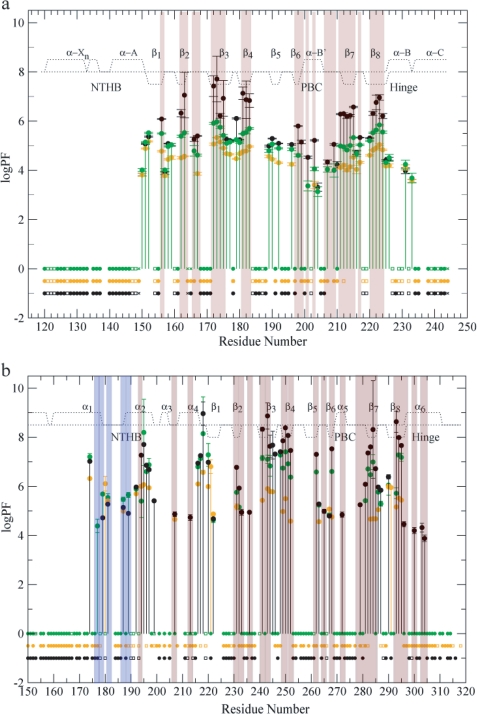
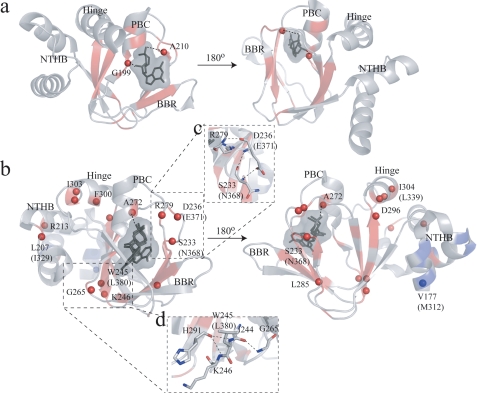
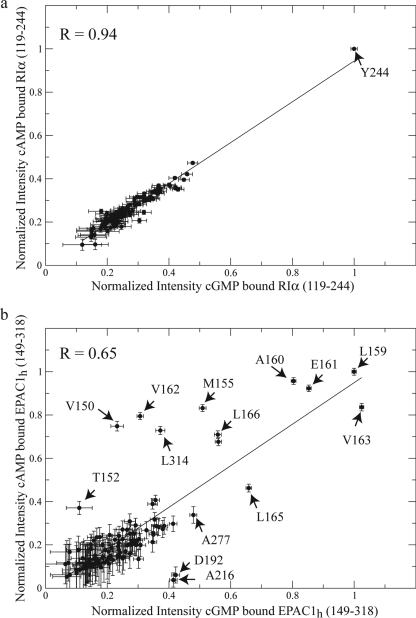
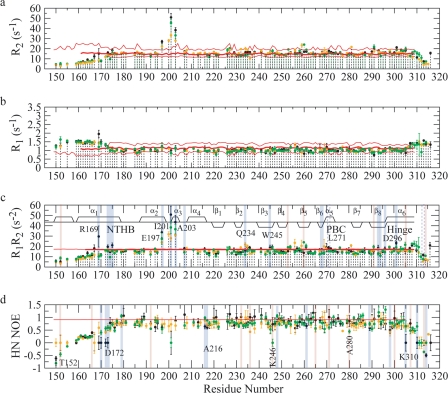
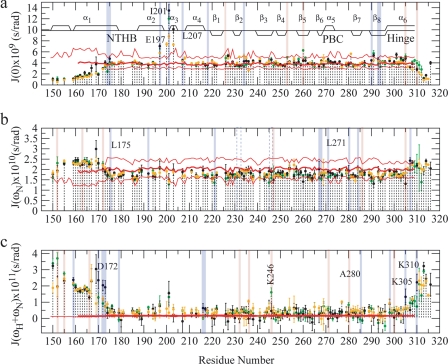
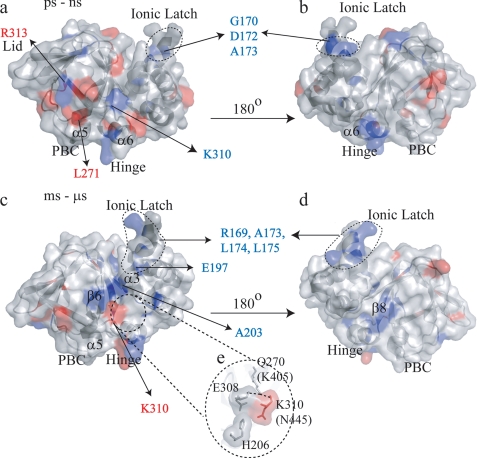
One of the mechanisms that minimize the aberrant cross-talk between cAMP- and cGMP-dependent signaling pathways relies on the selectivity of cAMP binding domains (CBDs). For instance, the CBDs of two critical eukaryotic cAMP receptors, i.e. protein kinase A (PKA) and the exchange protein activated by cAMP (EPAC), are both selectively activated by cAMP. However, the mechanisms underlying their cAMP versus cGMP selectivity are quite distinct. In PKA this selectivity is controlled mainly at the level of ligand affinity, whereas in EPAC it is mostly determined at the level of allostery. Currently, the molecular basis for these different selectivity mechanisms is not fully understood. We have therefore comparatively analyzed by NMR the cGMP-bound states of the essential CBDs of PKA and EPAC, revealing key differences between them. Specifically, cGMP binds PKA preserving the same syn base orientation as cAMP at the price of local steric clashes, which lead to a reduced affinity for cGMP. Unlike PKA, cGMP is recognized by EPAC in an anti conformation and generates several short and long range perturbations. Although these effects do not alter significantly the structure of the EPAC CBD investigated, remarkable differences in dynamics between the cAMP- and cGMP-bound states are detected for the ionic latch region. These observations suggest that one of the determinants of cGMP antagonism in EPAC is the modulation of the entropic control of inhibitory interactions and illustrate the pivotal role of allostery in determining signaling selectivity as a function of dynamic changes, even in the absence of significant affinity variations.
Figures
References
-
- Kim C., Vigil D., Anand G., Taylor S. S. (2006) Eur. J. Cell Biol. 85,651–654 - PubMed
-
- Rehmann H., Wittinghofer A., Bos J. L. (2007) Nat. Rev. Mol. Cell Biol. 8,63–73 - PubMed
-
- Kim C., Cheng C. Y., Saldanha S. A., Taylor S. S. (2007) Cell 130,1032–1043 - PubMed
-
- Pelligrino D. A., Wang Q. (1998) Prog. Neurobiol. 56,1–18 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
