

Mechanisms underlying short-term modulation of transmitter release by presynaptic depolarization
- PMID: 19403620
- PMCID: PMC2718256
- DOI: 10.1113/jphysiol.2009.168765
Mechanisms underlying short-term modulation of transmitter release by presynaptic depolarization
Abstract
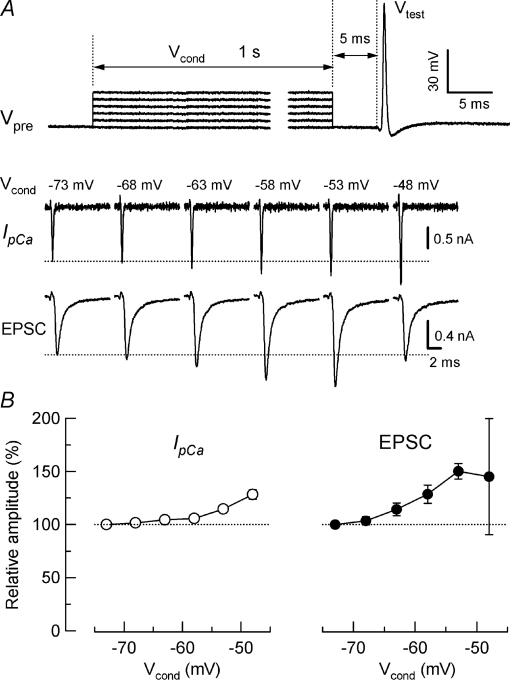
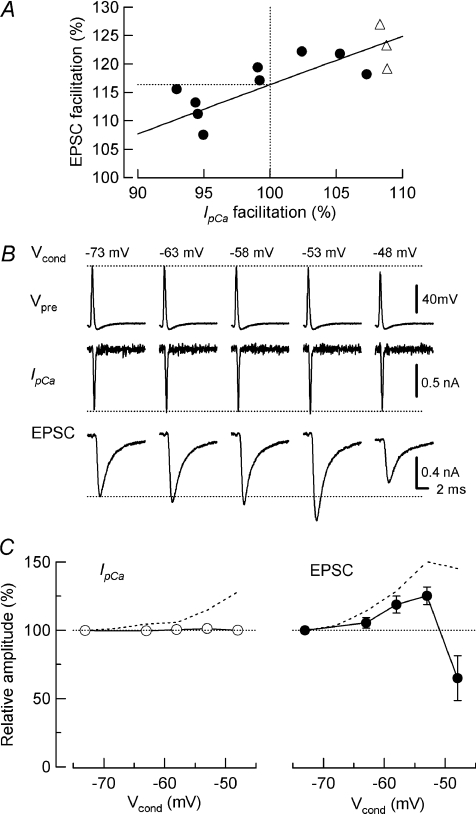
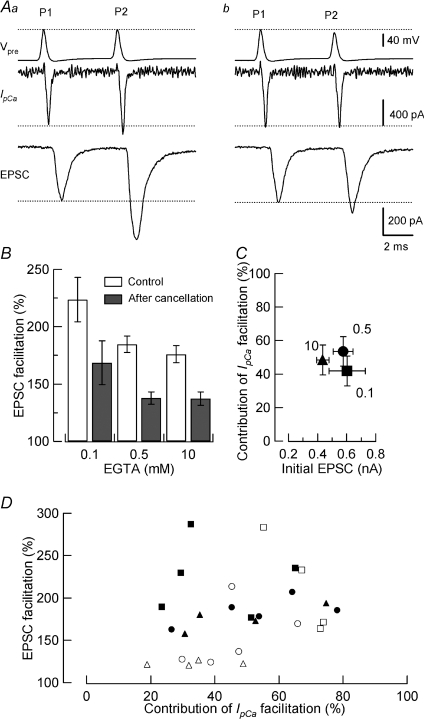
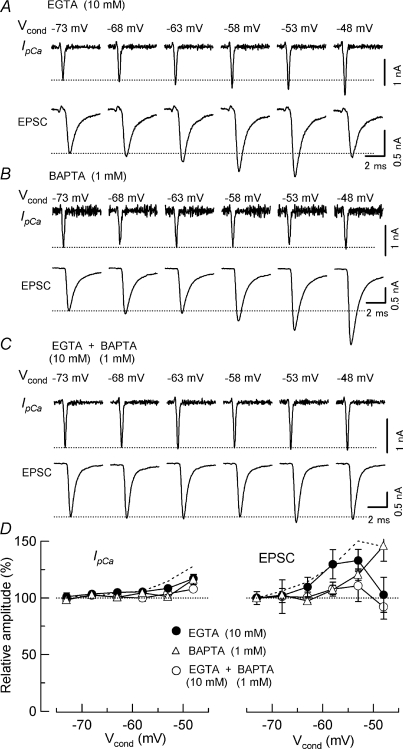
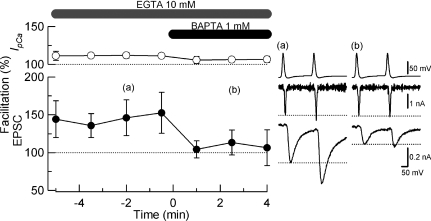
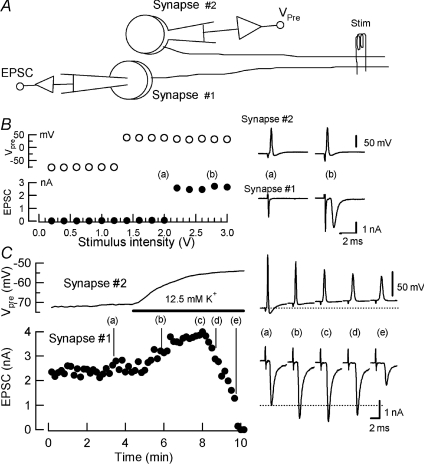
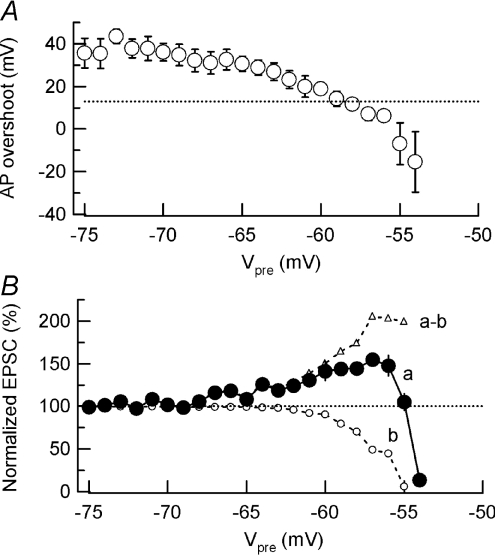
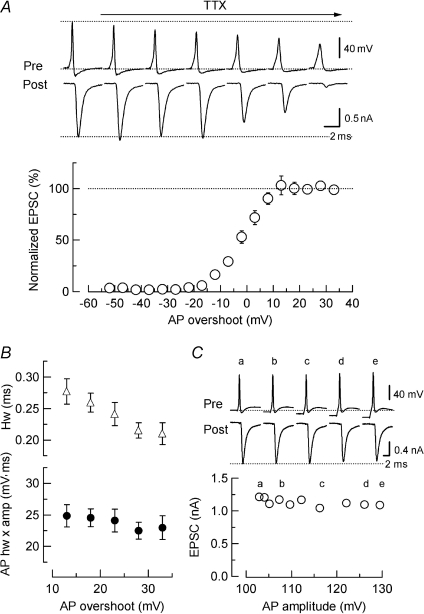
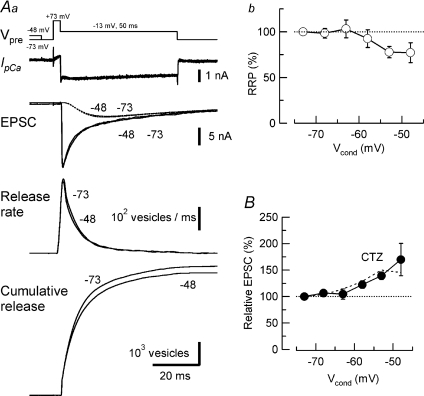
Presynaptic terminal depolarization modulates the efficacy of transmitter release. Residual Ca2+ remaining after presynaptic depolarization is thought to play a critical role in facilitation of transmitter release, but its downstream mechanism remains unclear. By making simultaneous pre- and postsynaptic recordings at the rodent calyx of Held synapse, we have investigated mechanisms involved in the facilitation and depression of postsynaptic currents induced by presynaptic depolarization. In voltage-clamp experiments, cancellation of the Ca2+-dependent presynaptic Ca2+ current (I(pCa)) facilitation revealed that this mechanism can account for 50% of postsynaptic current facilitation, irrespective of intraterminal EGTA concentrations. Intraterminal EGTA, loaded at 10 mM, failed to block postsynaptic current facilitation, but additional BAPTA at 1 mM abolished it. Potassium-induced sustained depolarization of non-dialysed presynaptic terminals caused a facilitation of postsynaptic currents, superimposed on a depression, with the latter resulting from reductions in presynaptic action potential amplitude and number of releasable vesicles. We conclude that presynaptic depolarization bidirectionally modulates transmitter release, and that the residual Ca2+ mechanism for synaptic facilitation operates in the immediate vicinity of voltage-gated Ca2+ channels in the nerve terminal.
Figures
References
-
- Alle H, Geiger JRP. Combined analog and action potential coding in hippocampal mossy fibers. Science. 2006;311:1290–1293. - PubMed
-
- Awatramani GB, Price GD, Trussell LO. Modulation of transmitter release by presynaptic resting potential and background calcium levels. Neuron. 2005;48:109–121. - PubMed
-
- Bertram R, Sherman A, Stanley EF. Singledomain/bound calcium hypothesis of transmitter release and facilitation. J Neurophysiol. 1996;75:1919–1931. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous