

Theiler's murine encephalomyelitis virus leader protein is the only nonstructural protein tested that induces apoptosis when transfected into mammalian cells
- PMID: 19403676
- PMCID: PMC2698581
- DOI: 10.1128/JVI.00353-09
Theiler's murine encephalomyelitis virus leader protein is the only nonstructural protein tested that induces apoptosis when transfected into mammalian cells
Abstract
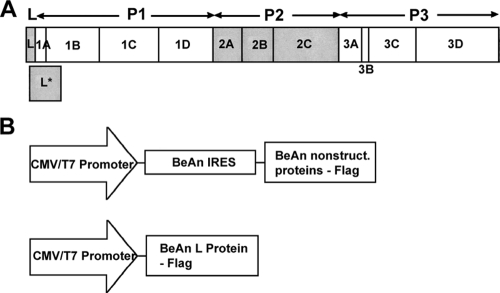
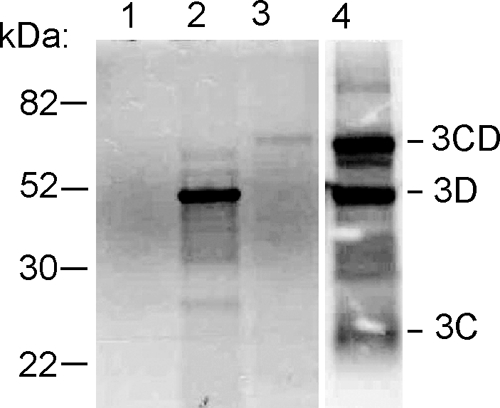
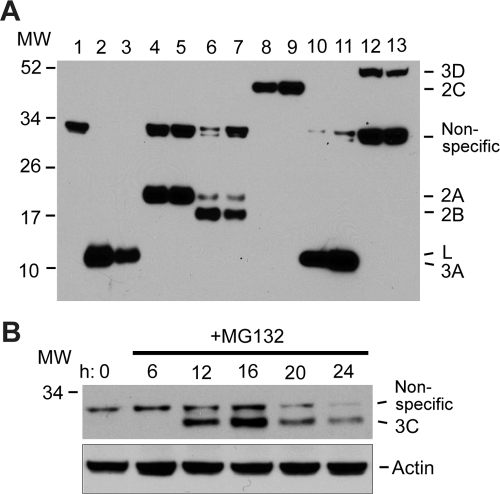
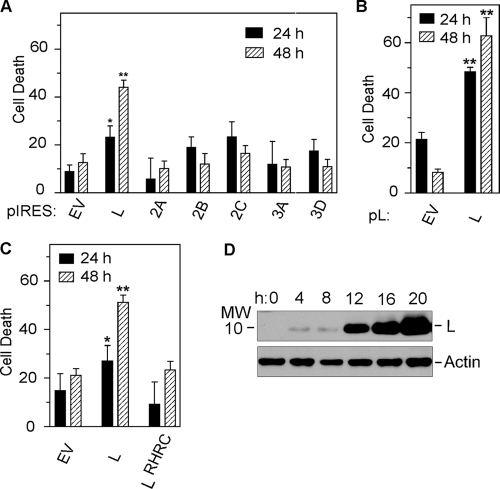
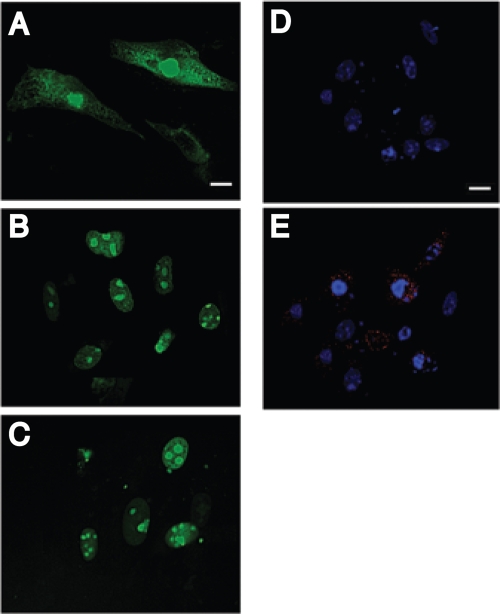
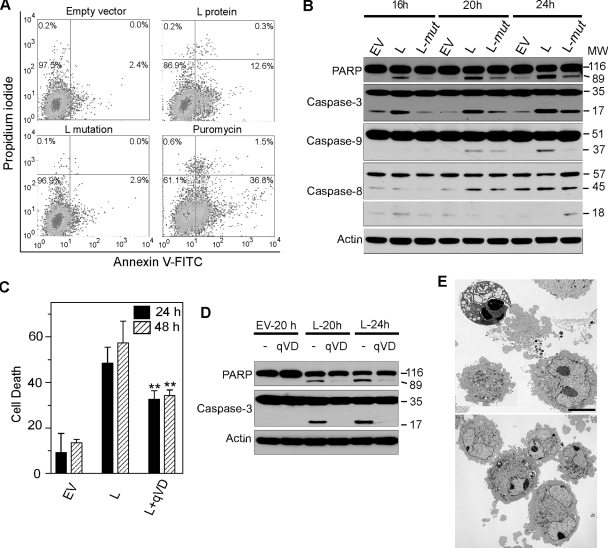
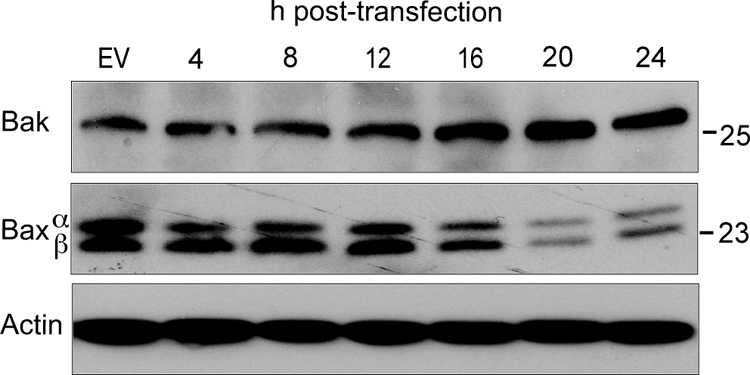
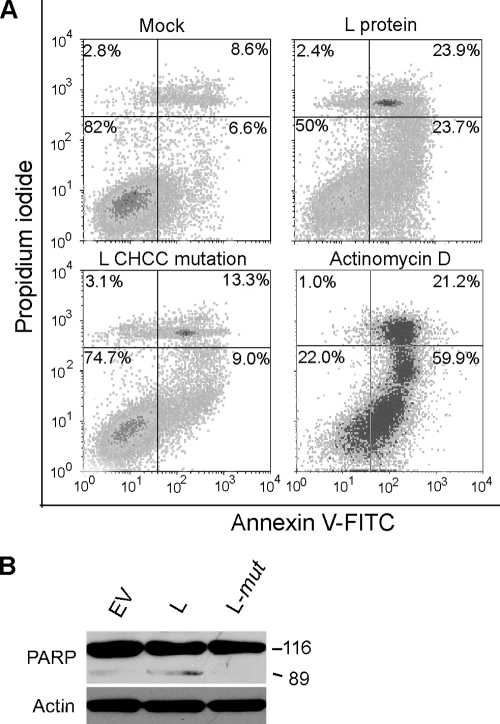
Theiler's murine encephalomyelitis virus (TMEV) induces two distinct cell death programs, necrosis and apoptosis. The apoptotic pathway is of particular interest because TMEV persists in the central nervous system of mice, largely in infiltrating macrophages, which undergo apoptosis. Infection of murine macrophages in culture induces apoptosis that is Bax dependent through the intrinsic or mitochondrial pathway, restricting infectious-virus yields and raising the possibility that apoptosis represents a mechanism to attenuate TMEV yet promote macrophage-to-macrophage spread during persistent infection. To help define the cellular stressors and upstream signaling events leading to apoptosis during TMEV infection, we screened baby hamster kidney (BHK-21) cells transfected to express individual nonstructural genes (except 3B) of the low-neurovirulence BeAn virus strain for cell death. Only expression of the leader protein led to apoptosis, as assessed by fluorescence-activated cell sorting analysis of propidium iodide- and annexin V-stained transfected cells, immunoblot analysis of poly(ADP-ribose) polymerase and caspase cleavages, electron microscopy, and inhibition of apoptosis by the pancaspase inhibitor qVD-OPh. After transfection, Bak and not Bax expression increased, suggesting that the apical pathway leading to activation of these Bcl-2 multi-BH-domain proapoptotic proteins differs in BeAn virus infection versus L transfection. Mutation to remove the CHCC Zn finger motif from L, a motif required by L to mediate inhibition of nucleocytoplasmic trafficking, significantly reduced L-protein-induced apoptosis in both BHK-21 and M1-D macrophages.
Figures
References
-
- Aminev, A. G., S. P. Amineva, and A. C. Palmenberg. 2003. Encephalomyocarditis virus (EMCV) proteins 2A and 2BCD localize to nuclei and inhibit cellular mRNA transcription but not rRNA transcription. Virus Res. 9559-73. - PubMed
-
- Amineva, S. P., A. G. Aminev, A. C. Palmenberg, and J. E. Gern. 2004. Rhinovirus 3C protease precursors 3CD and 3CD' localize to the nuclei of infected cells. J. Gen. Virol. 852969-2979. - PubMed
-
- Barco, A., E. Feduchi, and L. Carrasco. 2000. Poliovirus protease 3Cpro kills cells by apoptosis. Virology 266352-360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
