

Protracted withdrawal from alcohol and drugs of abuse impairs long-term potentiation of intrinsic excitability in the juxtacapsular bed nucleus of the stria terminalis
- PMID: 19403807
- PMCID: PMC2938175
- DOI: 10.1523/JNEUROSCI.5129-08.2009
Protracted withdrawal from alcohol and drugs of abuse impairs long-term potentiation of intrinsic excitability in the juxtacapsular bed nucleus of the stria terminalis
Abstract
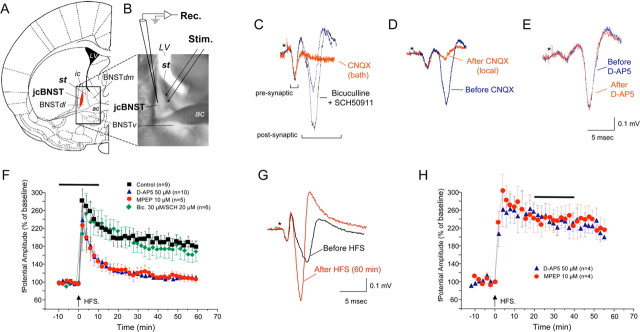
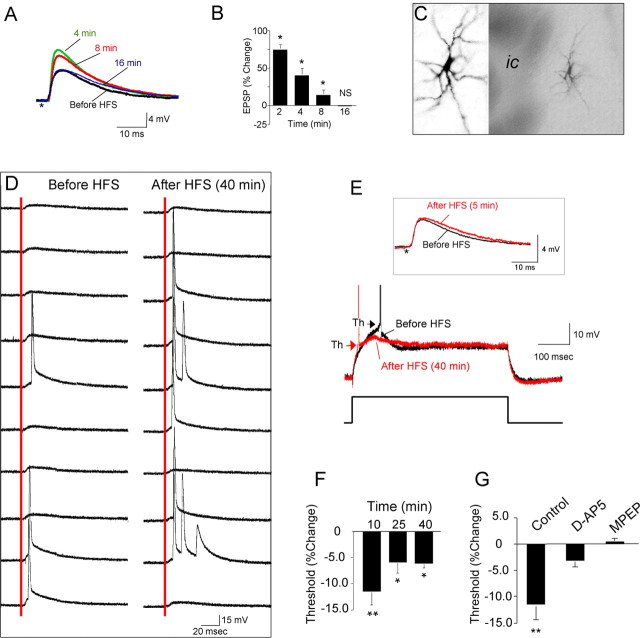
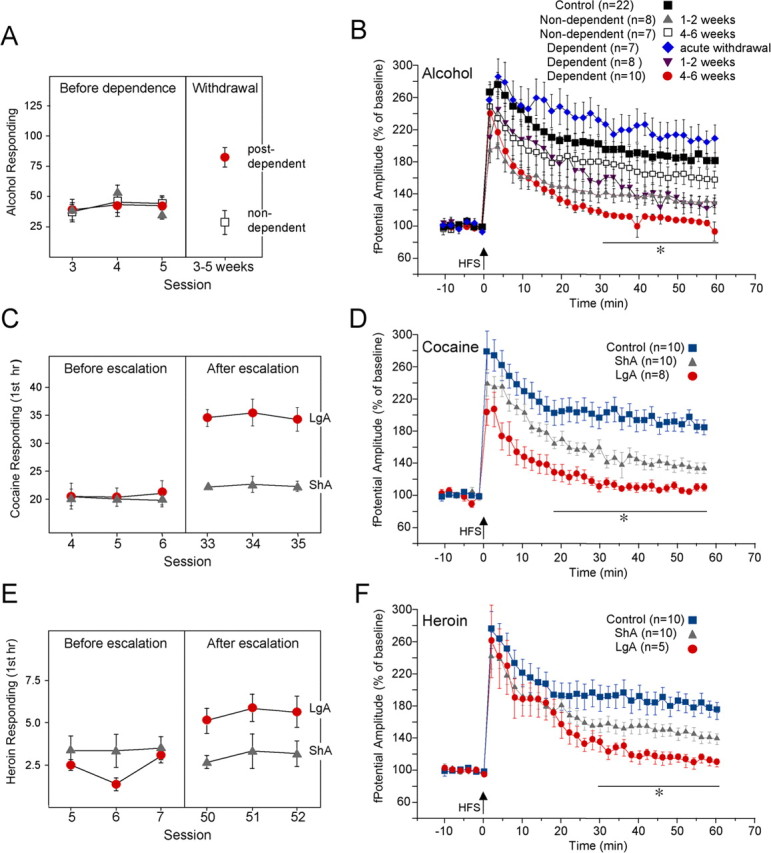
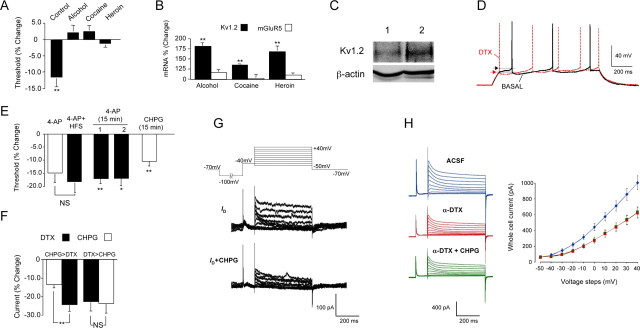
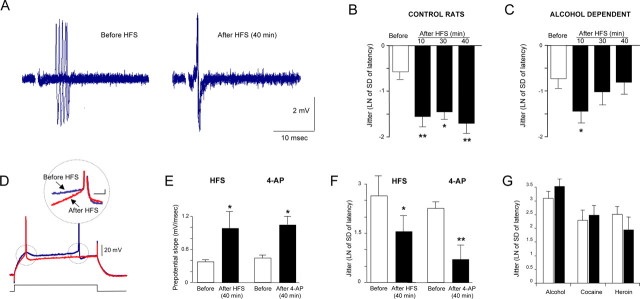
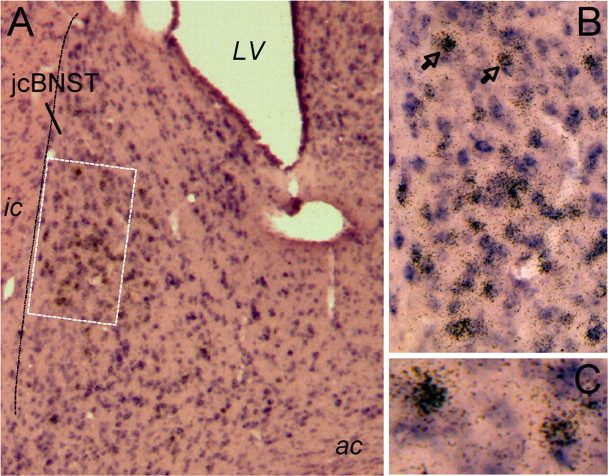
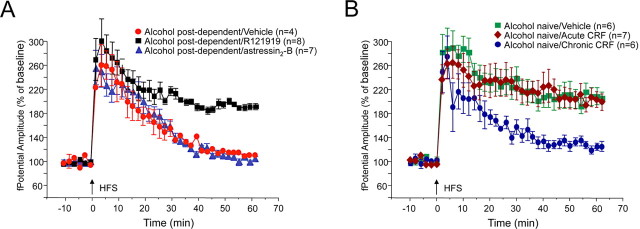
The juxtacapsular bed nucleus of the stria terminalis (jcBNST) is activated in response to basolateral amygdala (BLA) inputs through the stria terminalis and projects back to the anterior BLA and to the central nucleus of the amygdala. Here we show a form of long-term potentiation of the intrinsic excitability (LTP-IE) of jcBNST neurons in response to high-frequency stimulation of the stria terminalis. This LTP-IE, which was characterized by a decrease in the firing threshold and increased temporal fidelity of firing, was impaired during protracted withdrawal from self-administration of alcohol, cocaine, and heroin. Such impairment was graded and was more pronounced in rats that self-administered amounts of the drugs sufficient to maintain dependence. Dysregulation of the corticotropin-releasing factor (CRF) system has been implicated in manifestation of protracted withdrawal from dependent drug use. Administration of the selective corticotropin-releasing factor receptor 1 (CRF(1)) antagonist R121919 [2,5-dimethyl-3-(6-dimethyl-4-methylpyridin-3-yl)-7-dipropylamino-pyrazolo[1,5-a]pyrimidine)], but not of the CRF(2) antagonist astressin(2)-B, normalized jcBNST LTP-IE in animals with a history of alcohol dependence; repeated, but not acute, administration of CRF itself produced a decreased jcBNST LTP-IE. Thus, changes in the intrinsic properties of jcBNST neurons mediated by chronic activation of the CRF system may contribute to the persistent emotional dysregulation associated with protracted withdrawal.
Figures
Similar articles
-
Intrinsic neuronal plasticity in the juxtacapsular nucleus of the bed nuclei of the stria terminalis (jcBNST).Prog Neuropsychopharmacol Biol Psychiatry. 2009 Nov 13;33(8):1347-55. doi: 10.1016/j.pnpbp.2009.08.003. Epub 2009 Aug 12. Prog Neuropsychopharmacol Biol Psychiatry. 2009. PMID: 19683025 Free PMC article.
-
Opiate dependence induces cell type-specific plasticity of intrinsic membrane properties in the rat juxtacapsular bed nucleus of stria terminalis (jcBNST).Psychopharmacology (Berl). 2017 Dec;234(23-24):3485-3498. doi: 10.1007/s00213-017-4732-4. Epub 2017 Oct 6. Psychopharmacology (Berl). 2017. PMID: 28986608 Free PMC article.
-
Excitability of jcBNST neurons is reduced in alcohol-dependent animals during protracted alcohol withdrawal.PLoS One. 2012;7(8):e42313. doi: 10.1371/journal.pone.0042313. Epub 2012 Aug 21. PLoS One. 2012. PMID: 22927925 Free PMC article.
-
Role of the bed nucleus of the stria terminalis versus the amygdala in fear, stress, and anxiety.Eur J Pharmacol. 2003 Feb 28;463(1-3):199-216. doi: 10.1016/s0014-2999(03)01282-2. Eur J Pharmacol. 2003. PMID: 12600711 Review.
-
Noradrenergic transmission in the extended amygdala: role in increased drug-seeking and relapse during protracted drug abstinence.Brain Struct Funct. 2008 Sep;213(1-2):43-61. doi: 10.1007/s00429-008-0191-3. Epub 2008 Jul 24. Brain Struct Funct. 2008. PMID: 18651175 Free PMC article. Review.
Cited by
-
Corticotropin-Releasing Factor (CRF) Neurocircuitry and Neuropharmacology in Alcohol Drinking.Handb Exp Pharmacol. 2018;248:435-471. doi: 10.1007/164_2017_86. Handb Exp Pharmacol. 2018. PMID: 29374836 Free PMC article. Review.
-
Conventional measures of intrinsic excitability are poor estimators of neuronal activity under realistic synaptic inputs.PLoS Comput Biol. 2021 Sep 16;17(9):e1009378. doi: 10.1371/journal.pcbi.1009378. eCollection 2021 Sep. PLoS Comput Biol. 2021. PMID: 34529674 Free PMC article.
-
Distinct forms of Gq-receptor-dependent plasticity of excitatory transmission in the BNST are differentially affected by stress.Proc Natl Acad Sci U S A. 2010 Feb 2;107(5):2271-6. doi: 10.1073/pnas.0905568107. Epub 2010 Jan 19. Proc Natl Acad Sci U S A. 2010. PMID: 20133871 Free PMC article.
-
Corticotropin releasing factor and drug seeking in substance use disorders: Preclinical evidence and translational limitations.Addict Neurosci. 2022 Dec;4:100038. doi: 10.1016/j.addicn.2022.100038. Epub 2022 Oct 1. Addict Neurosci. 2022. PMID: 36531188 Free PMC article.
-
CRF modulation of central monoaminergic function: Implications for sex differences in alcohol drinking and anxiety.Alcohol. 2018 Nov;72:33-47. doi: 10.1016/j.alcohol.2018.01.007. Epub 2018 Feb 2. Alcohol. 2018. PMID: 30217435 Free PMC article. Review.
References
-
- Ahmed SH, Kenny PJ, Koob GF, Markou A. Neurobiological evidence for hedonic allostasis associated with escalating cocaine use. Nat Neurosci. 2002;5:625–626. - PubMed
-
- Aizenman CD, Linden DJ. Rapid, synaptically driven increases in the intrinsic excitability of cerebellar deep nuclear neurons. Nat Neurosci. 2000;3:109–111. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P50 AA006420/AA/NIAAA NIH HHS/United States
- R37 AA008459/AA/NIAAA NIH HHS/United States
- AA016587/AA/NIAAA NIH HHS/United States
- R01 AA016587/AA/NIAAA NIH HHS/United States
- AA013191/AA/NIAAA NIH HHS/United States
- ImNIH/Intramural NIH HHS/United States
- R01 DA004398/DA/NIDA NIH HHS/United States
- R01 AA013191/AA/NIAAA NIH HHS/United States
- DA013821/DA/NIDA NIH HHS/United States
- R01 DA013821/DA/NIDA NIH HHS/United States
- AA006420/AA/NIAAA NIH HHS/United States
- R01 AA008459/AA/NIAAA NIH HHS/United States
- DA004398/DA/NIDA NIH HHS/United States
- AA008459/AA/NIAAA NIH HHS/United States
- R01 DA004043/DA/NIDA NIH HHS/United States
LinkOut - more resources
Full Text Sources