

p75 and TrkA signaling regulates sympathetic neuronal firing patterns via differential modulation of voltage-gated currents
- PMID: 19403809
- PMCID: PMC3326291
- DOI: 10.1523/JNEUROSCI.3503-08.2009
p75 and TrkA signaling regulates sympathetic neuronal firing patterns via differential modulation of voltage-gated currents
Abstract
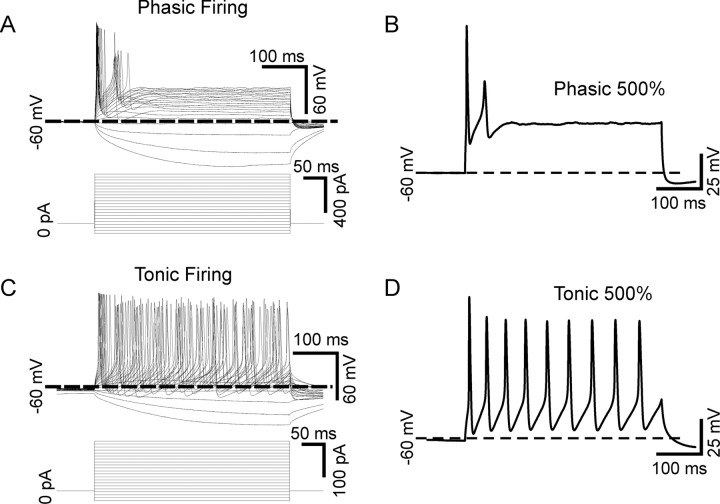
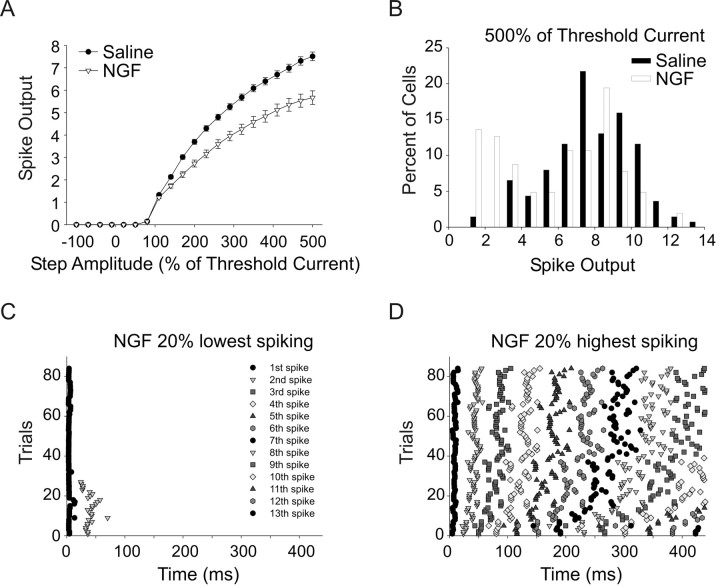
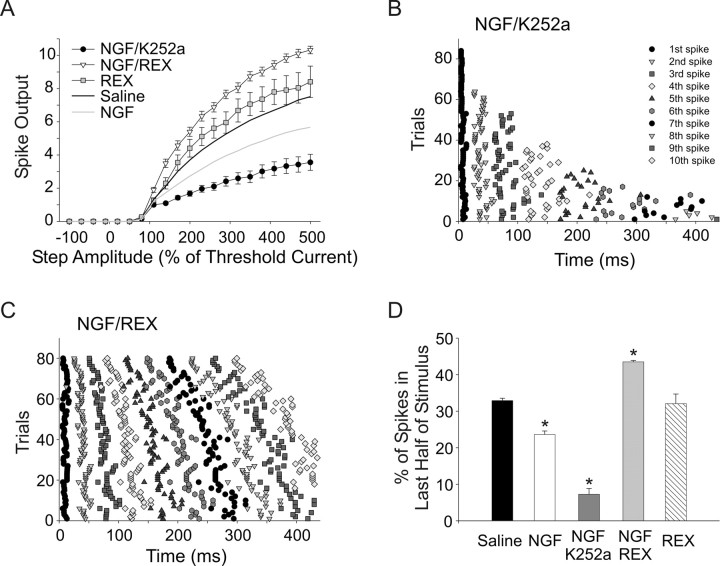
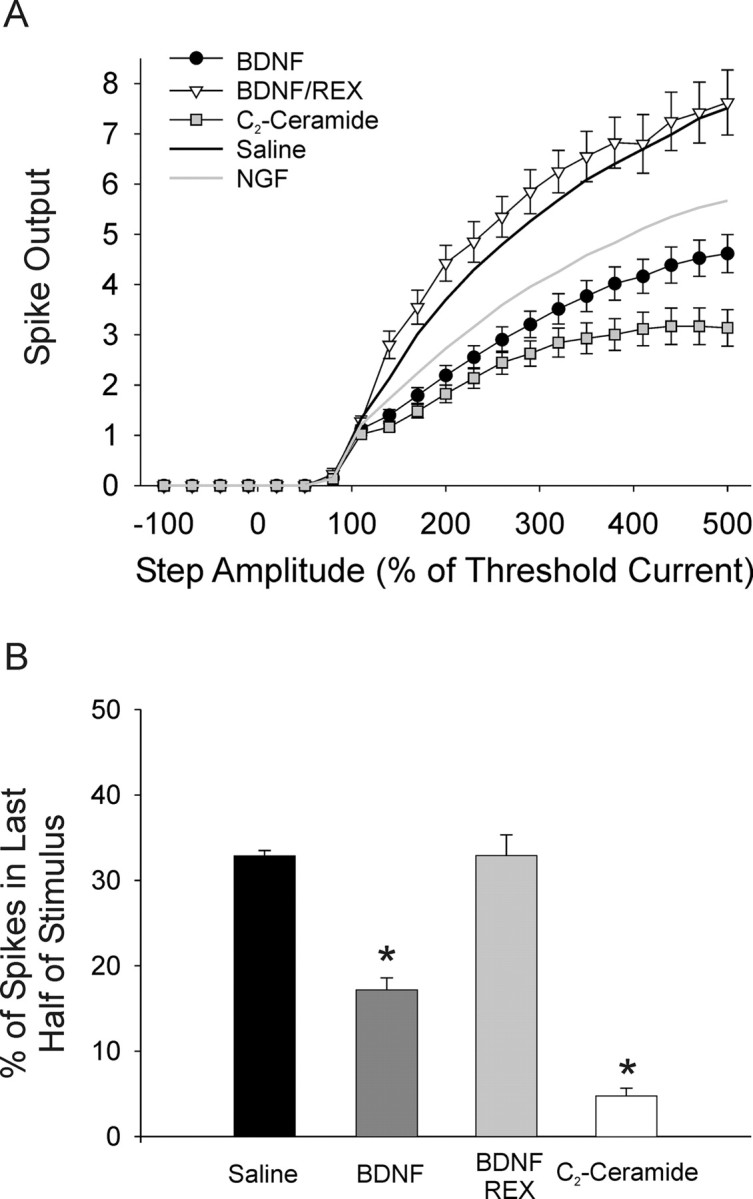
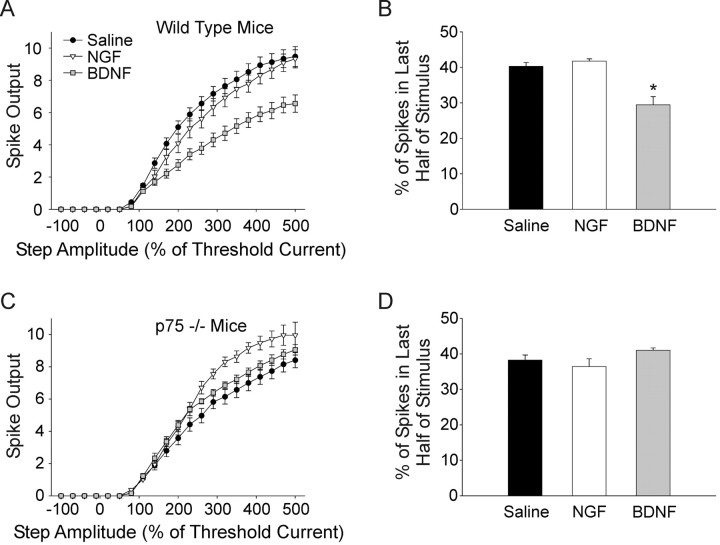
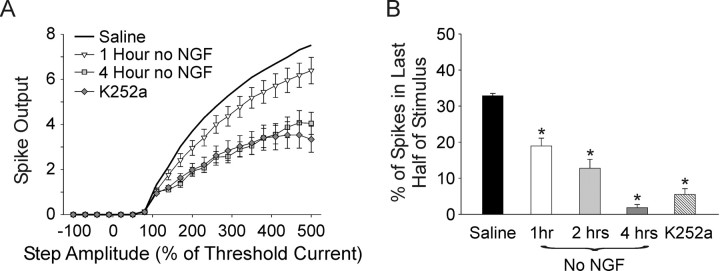
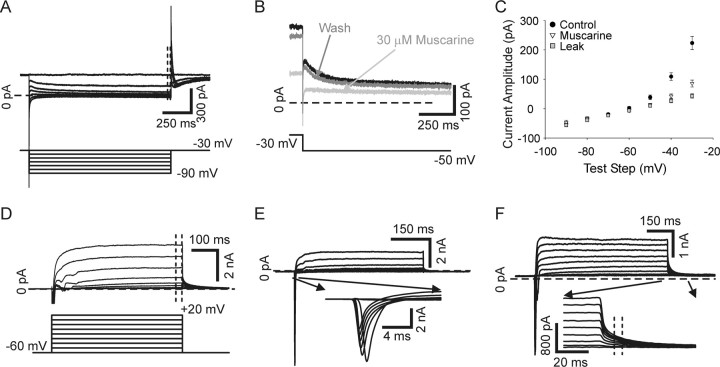
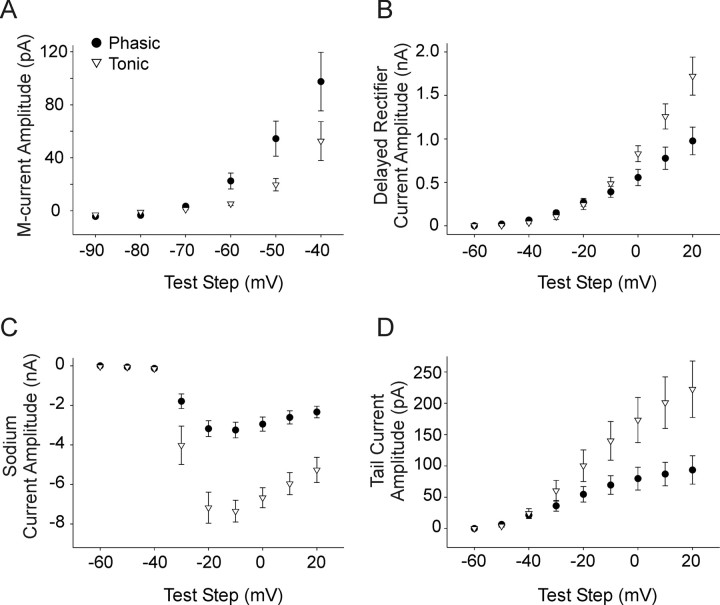
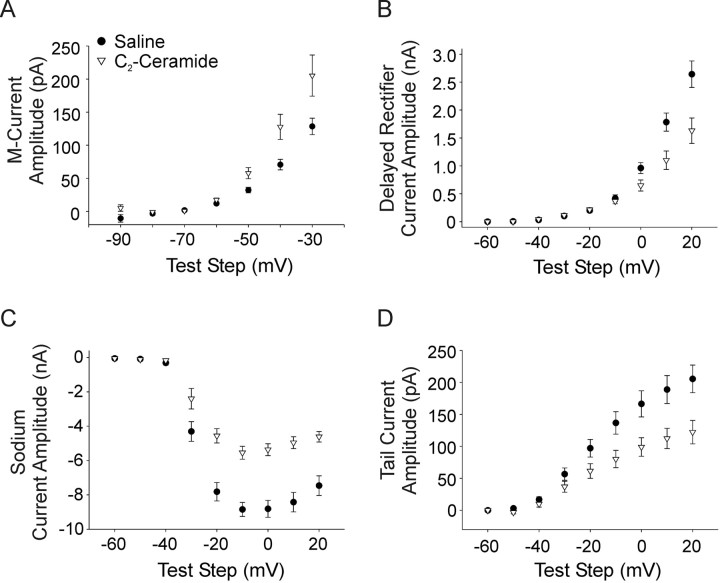
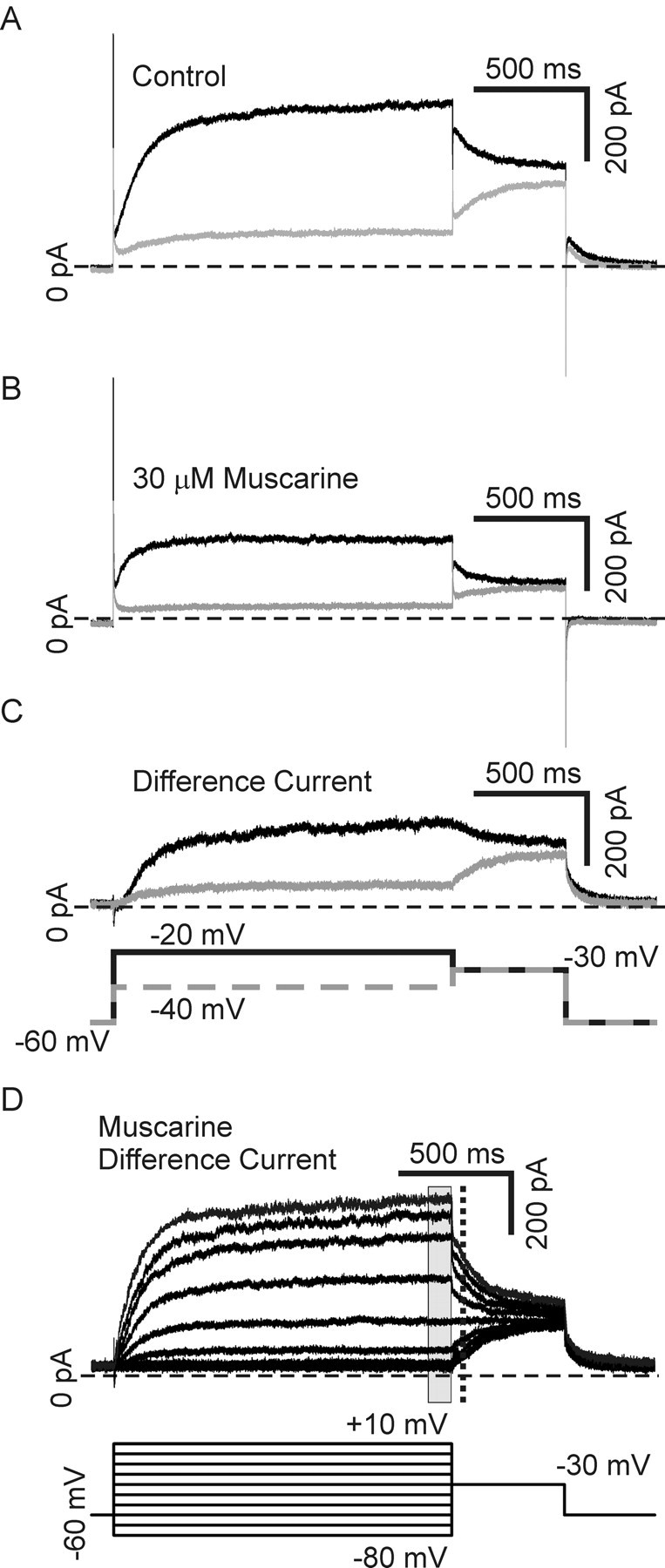
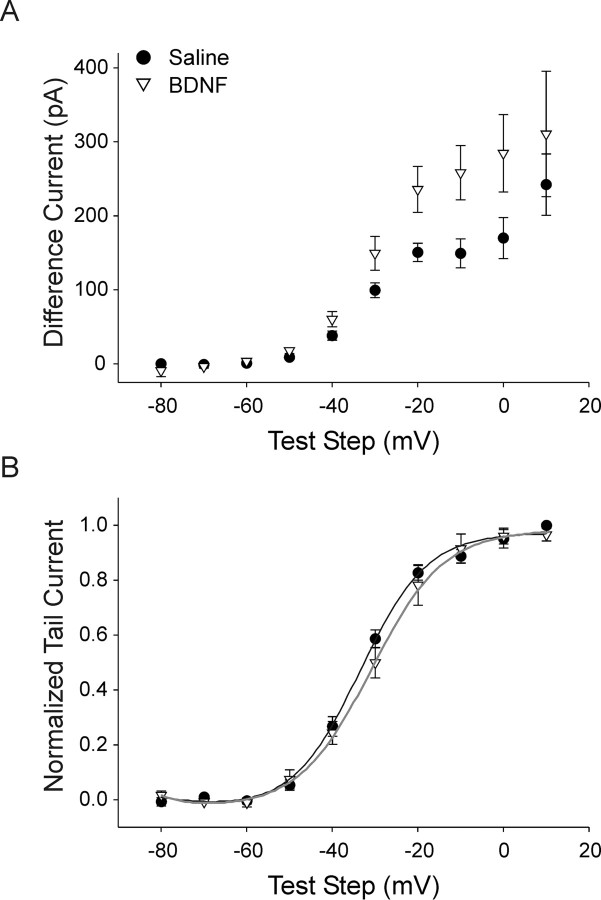
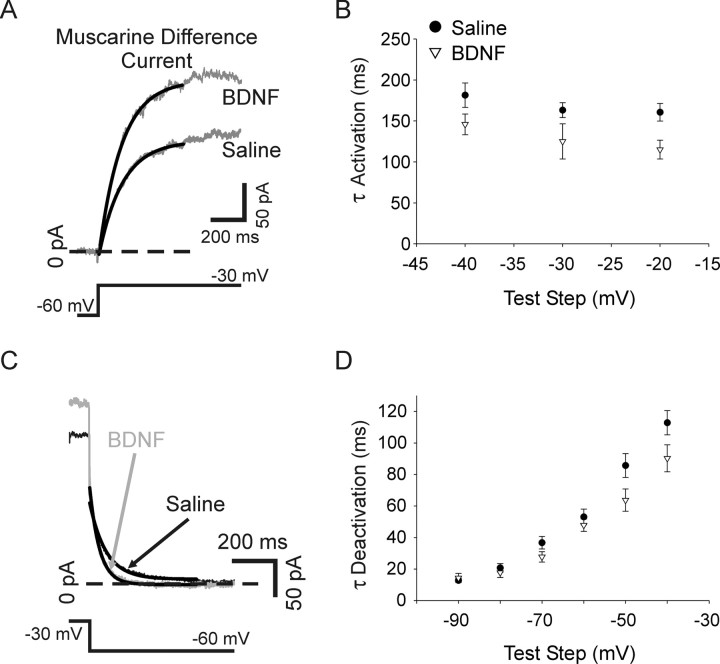
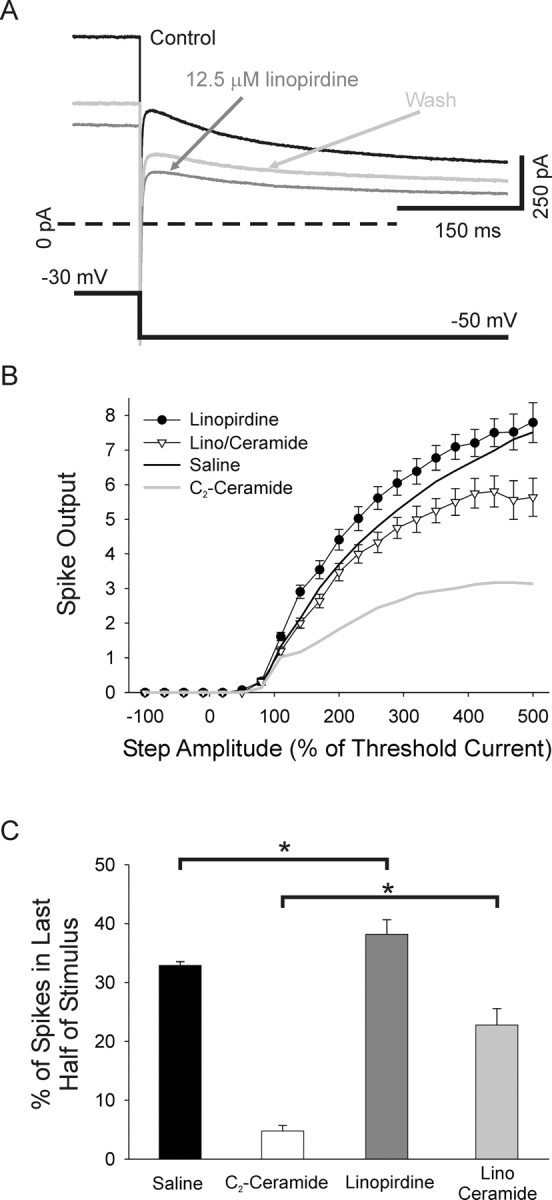
Neurotrophins such as nerve growth factor (NGF) and brain-derived neurotrophic factor (BDNF) act through the tropomyosin-related receptor tyrosine kinases (Trk) and the pan-neurotrophin receptor (p75) to regulate complex developmental and functional properties of neurons. While NGF activates both receptor types in sympathetic neurons, differential signaling through TrkA and p75 can result in widely divergent functional outputs for neuronal survival, growth, and synaptic function. Here we show that TrkA and p75 signaling pathways have opposing effects on the firing properties of sympathetic neurons, and define a mechanism whereby the relative level of signaling through these two receptors sets firing patterns via coordinate regulation of a set of ionic currents. We show that signaling through the p75 pathway causes sympathetic neurons to fire in a phasic pattern showing marked accommodation. Signaling through the NGF-specific TrkA, on the other hand, causes cells to fire tonically. Neurons switch rapidly between firing patterns, on the order of minutes to hours. We show that changes in firing patterns are caused by neurotrophin-dependent regulation of at least four voltage-gated currents: the sodium current and the M-type, delayed rectifier, and calcium-dependent potassium currents. Neurotrophin release, and thus receptor activation, varies among somatic tissues and physiological state. Thus, these data suggest that target-derived neurotrophins may be an important determinant of the characteristic electrical properties of sympathetic neurons and therefore regulate the functional output of the sympathetic nervous system.
Figures
Similar articles
-
Neurotrophins and target interactions in the development and regulation of sympathetic neuron electrical and synaptic properties.Auton Neurosci. 2009 Nov 17;151(1):46-60. doi: 10.1016/j.autneu.2009.08.009. Epub 2009 Sep 13. Auton Neurosci. 2009. PMID: 19748836 Free PMC article. Review.
-
Nerve growth factor regulates substance P in adult sensory neurons through both TrkA and p75 receptors.Exp Neurol. 2006 Feb;197(2):430-6. doi: 10.1016/j.expneurol.2005.10.006. Epub 2005 Nov 21. Exp Neurol. 2006. PMID: 16300761
-
Differential dependency of unmyelinated and A delta epidermal and upper dermal innervation on neurotrophins, trk receptors, and p75LNGFR.Dev Biol. 1998 Jun 1;198(1):57-81. Dev Biol. 1998. PMID: 9640332
-
Differential modulation of neuron survival during development by nerve growth factor binding to the p75 neurotrophin receptor.J Biol Chem. 1997 Jun 27;272(26):16322-8. doi: 10.1074/jbc.272.26.16322. J Biol Chem. 1997. PMID: 9195937
-
The p75 neurotrophin receptor and neuronal apoptosis.Prog Neurobiol. 2000 Jun;61(2):205-29. doi: 10.1016/s0301-0082(99)00056-8. Prog Neurobiol. 2000. PMID: 10704998 Review.
Cited by
-
Ngfr+ cholinergic projection from SI/nBM to mPFC selectively regulates temporal order recognition memory.Nat Commun. 2024 Aug 26;15(1):7342. doi: 10.1038/s41467-024-51707-w. Nat Commun. 2024. PMID: 39187496 Free PMC article.
-
Neurotrophins and target interactions in the development and regulation of sympathetic neuron electrical and synaptic properties.Auton Neurosci. 2009 Nov 17;151(1):46-60. doi: 10.1016/j.autneu.2009.08.009. Epub 2009 Sep 13. Auton Neurosci. 2009. PMID: 19748836 Free PMC article. Review.
-
Effects of sustained proNGF blockade on attentional capacities in aged rats with compromised cholinergic system.Neuroscience. 2014 Mar 7;261:118-32. doi: 10.1016/j.neuroscience.2013.12.042. Epub 2013 Dec 27. Neuroscience. 2014. PMID: 24374328 Free PMC article.
-
A Calcium-Dependent Chloride Current Increases Repetitive Firing in Mouse Sympathetic Neurons.Front Physiol. 2018 May 14;9:508. doi: 10.3389/fphys.2018.00508. eCollection 2018. Front Physiol. 2018. PMID: 29867553 Free PMC article.
-
Extracellular pH and neuronal depolarization serve as dynamic switches to rapidly mobilize trkA to the membrane of adult sensory neurons.J Neurosci. 2013 May 8;33(19):8202-15. doi: 10.1523/JNEUROSCI.4408-12.2013. J Neurosci. 2013. PMID: 23658159 Free PMC article.
References
-
- Akopian AN, Souslova V, England S, Okuse K, Ogata N, Ure J, Smith A, Kerr BJ, McMahon SB, Boyce S, Hill R, Stanfa LC, Dickenson AH, Wood JN. The tetrodotoxin-resistant sodium channel SNS has a specialized function in pain pathways. Nat Neurosci. 1999;2:541–548. - PubMed
-
- Anderson RL, Jobling P, Gibbins IL. Development of electrophysiological and morphological diversity in autonomic neurons. J Neurophysiol. 2001;86:1237–1251. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials