

Mechanism of differential control of NMDA receptor activity by NR2 subunits
- PMID: 19404260
- PMCID: PMC2711440
- DOI: 10.1038/nature07993
Mechanism of differential control of NMDA receptor activity by NR2 subunits
Abstract
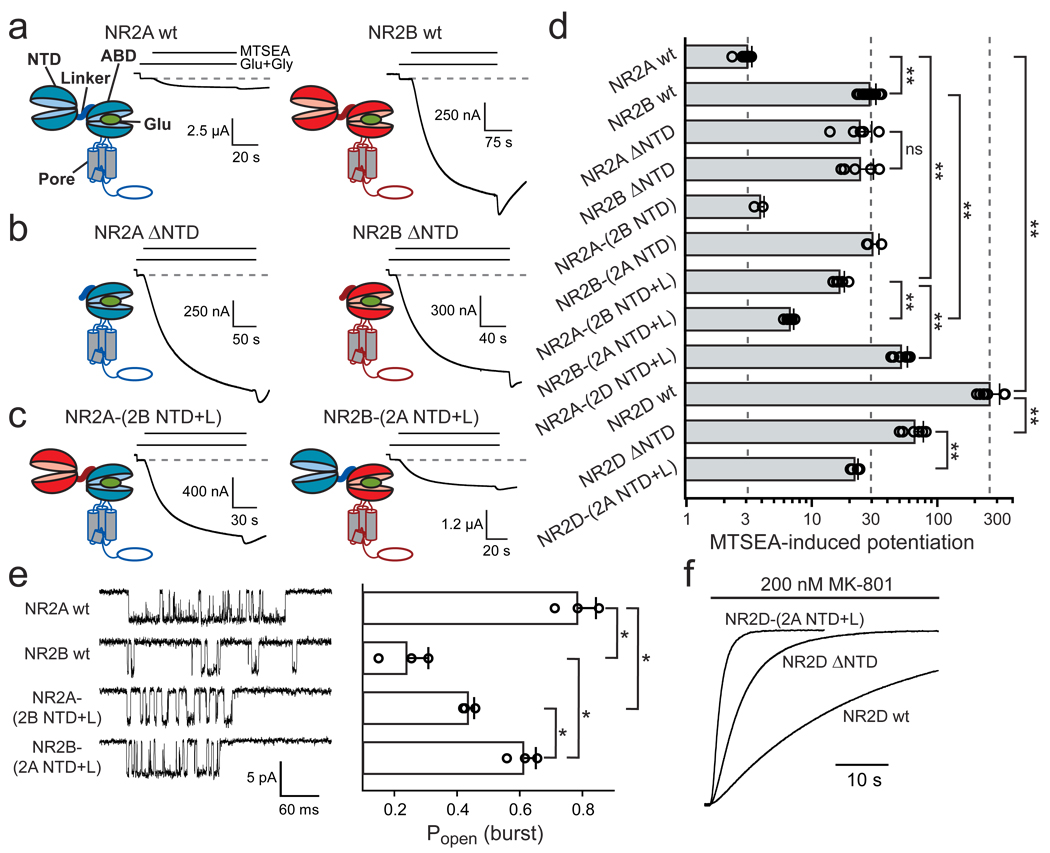
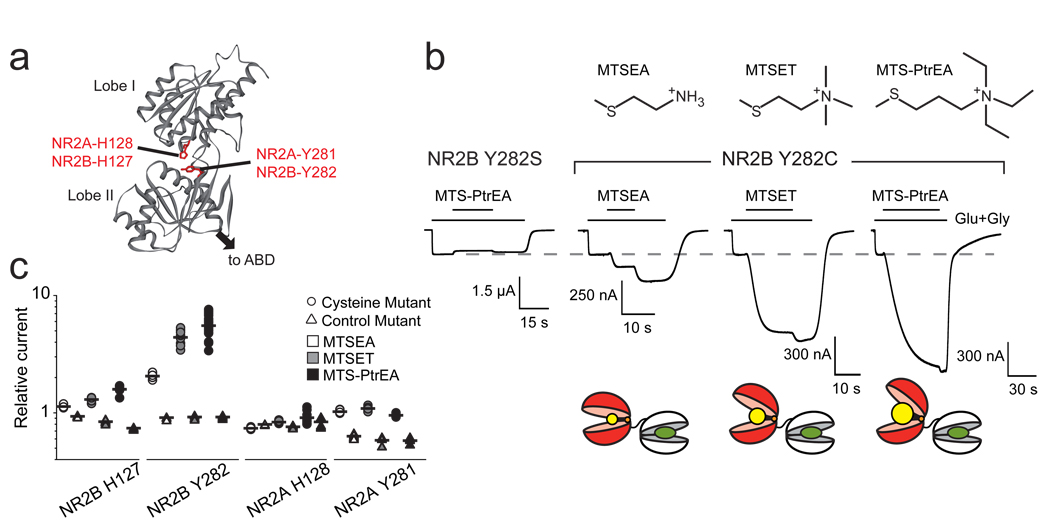
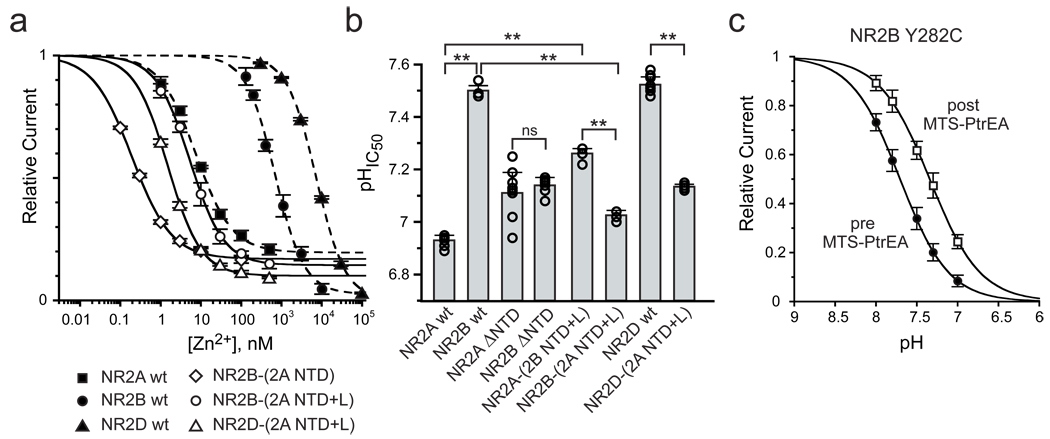
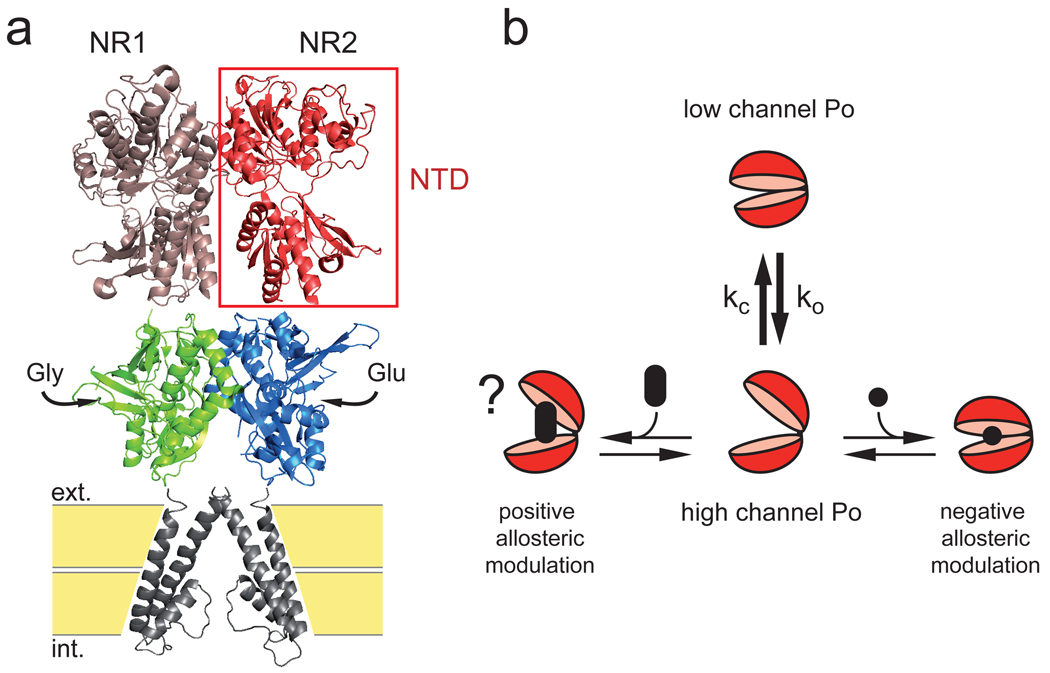
N-methyl-d-aspartate (NMDA) receptors (NMDARs) are a major class of excitatory neurotransmitter receptors in the central nervous system. They form glutamate-gated ion channels that are highly permeable to calcium and mediate activity-dependent synaptic plasticity. NMDAR dysfunction is implicated in multiple brain disorders, including stroke, chronic pain and schizophrenia. NMDARs exist as multiple subtypes with distinct pharmacological and biophysical properties that are largely determined by the type of NR2 subunit (NR2A to NR2D) incorporated in the heteromeric NR1/NR2 complex. A fundamental difference between NMDAR subtypes is their channel maximal open probability (P(o)), which spans a 50-fold range from about 0.5 for NR2A-containing receptors to about 0.01 for receptors containing NR2C and NR2D; NR2B-containing receptors have an intermediate value (about 0.1). These differences in P(o) confer unique charge transfer capacities and signalling properties on each receptor subtype. The molecular basis for this profound difference in activity between NMDAR subtypes is unknown. Here we show that the subunit-specific gating of NMDARs is controlled by the region formed by the NR2 amino-terminal domain (NTD), an extracellular clamshell-like domain previously shown to bind allosteric inhibitors, and the short linker connecting the NTD to the agonist-binding domain (ABD). The subtype specificity of NMDAR P(o) largely reflects differences in the spontaneous (ligand-independent) equilibrium between open-cleft and closed-cleft conformations of the NR2-NTD. This NTD-driven gating control also affects pharmacological properties by setting the sensitivity to the endogenous inhibitors zinc and protons. Our results provide a proof of concept for a drug-based bidirectional control of NMDAR activity by using molecules acting either as NR2-NTD 'closers' or 'openers' promoting receptor inhibition or potentiation, respectively.
Figures
References
-
- Dingledine R, Borges K, Bowie D, Traynelis SF. The glutamate receptor ion channels. Pharmacol Rev. 1999;51:7–61. - PubMed
-
- Kemp JA, McKernan RM. NMDA receptor pathways as drug targets. Nat Neurosci. 2002;5(Suppl):1039–1042. - PubMed
-
- Paoletti P, Neyton J. NMDA receptor subunits: function and pharmacology. Curr Opin Pharmacol. 2007;7:39–47. - PubMed
-
- Cull-Candy SG, Leszkiewicz DN. Role of distinct NMDA receptor subtypes at central synapses. Sci STKE. 2004;2004:re16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
