

Extracting kinematic parameters for monkey bipedal walking from cortical neuronal ensemble activity
- PMID: 19404411
- PMCID: PMC2659168
- DOI: 10.3389/neuro.07.003.2009
Extracting kinematic parameters for monkey bipedal walking from cortical neuronal ensemble activity
Abstract
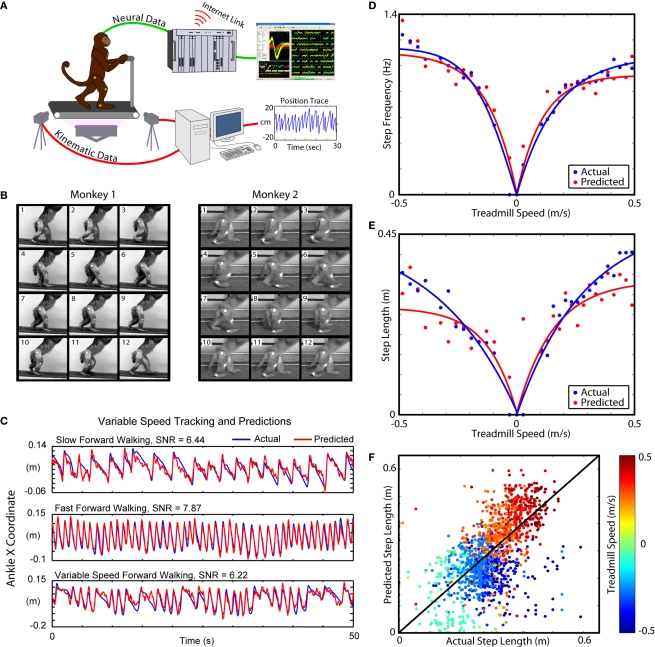
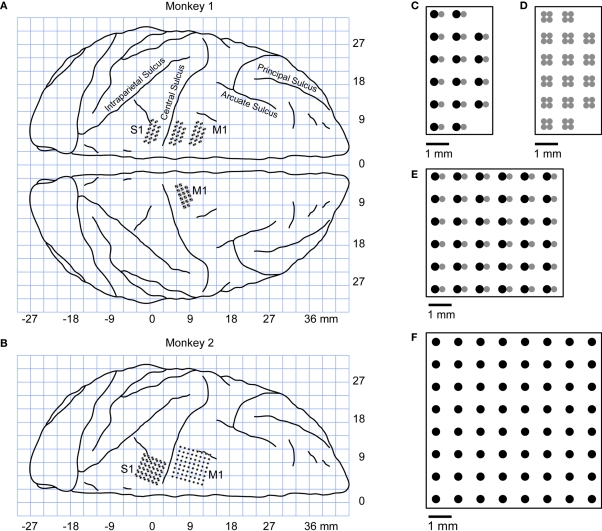
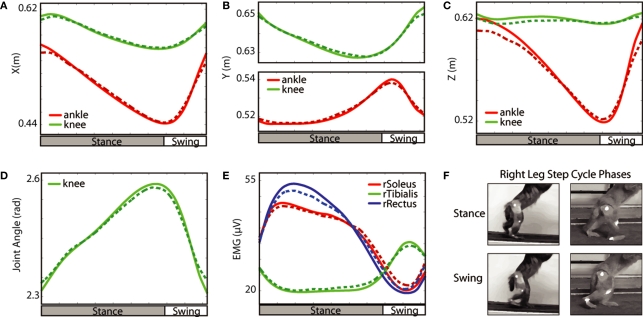
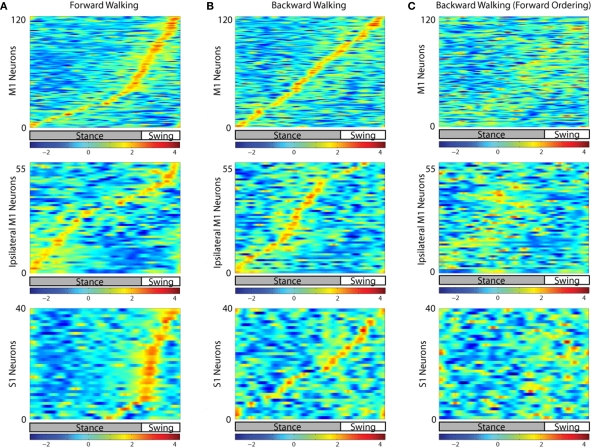
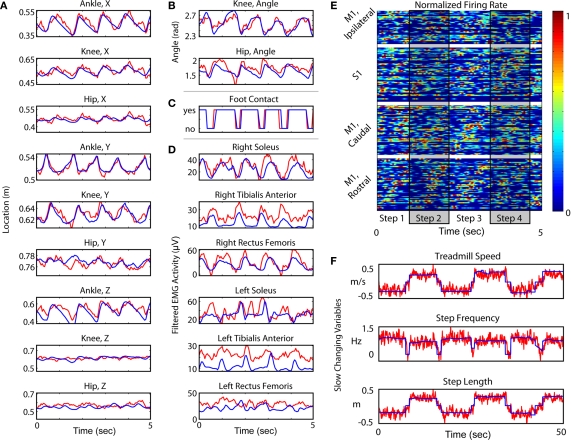
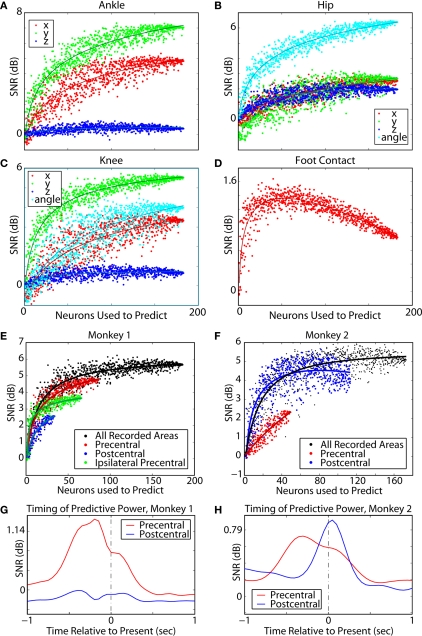
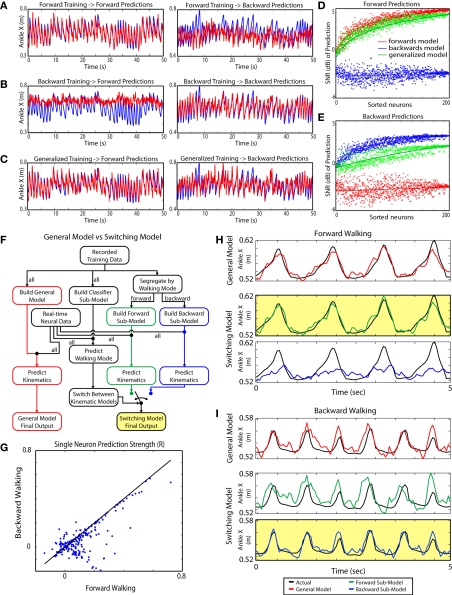
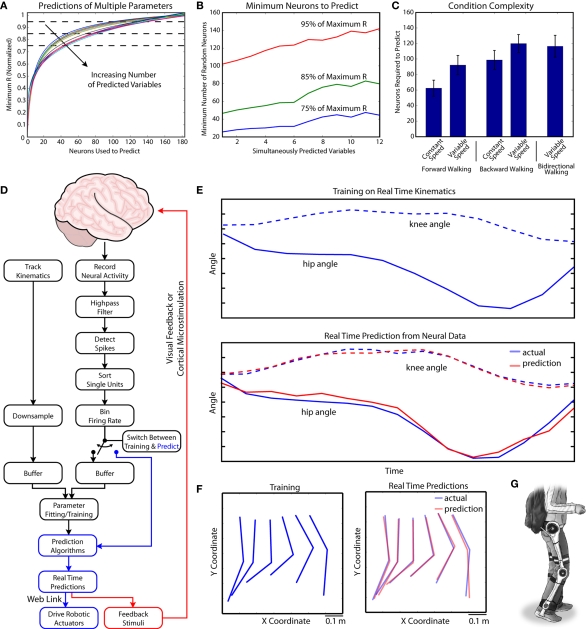
The ability to walk may be critically impacted as the result of neurological injury or disease. While recent advances in brain-machine interfaces (BMIs) have demonstrated the feasibility of upper-limb neuroprostheses, BMIs have not been evaluated as a means to restore walking. Here, we demonstrate that chronic recordings from ensembles of cortical neurons can be used to predict the kinematics of bipedal walking in rhesus macaques - both offline and in real time. Linear decoders extracted 3D coordinates of leg joints and leg muscle electromyograms from the activity of hundreds of cortical neurons. As more complex patterns of walking were produced by varying the gait speed and direction, larger neuronal populations were needed to accurately extract walking patterns. Extraction was further improved using a switching decoder which designated a submodel for each walking paradigm. We propose that BMIs may one day allow severely paralyzed patients to walk again.
Keywords: brain–machine interface; locomotion; neuronal ensemble recordings; neuroprosthetics; primate; sensorimotor.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials