

Characterization of the nucleation barriers for protein aggregation and amyloid formation
- PMID: 19404419
- PMCID: PMC2639838
- DOI: 10.2976/1.2760023
Characterization of the nucleation barriers for protein aggregation and amyloid formation
Abstract
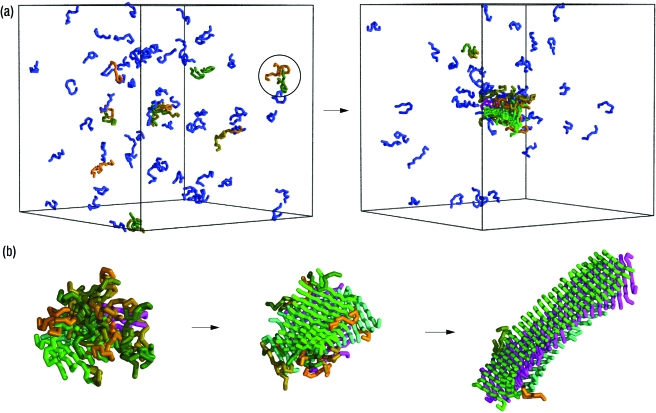
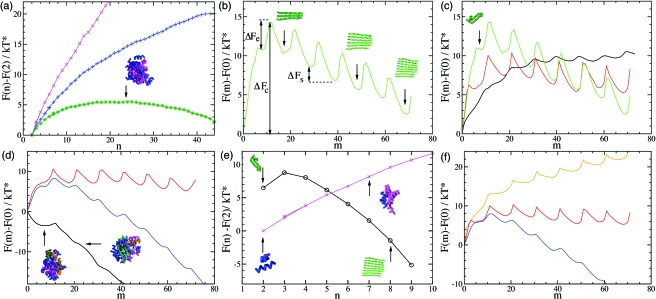
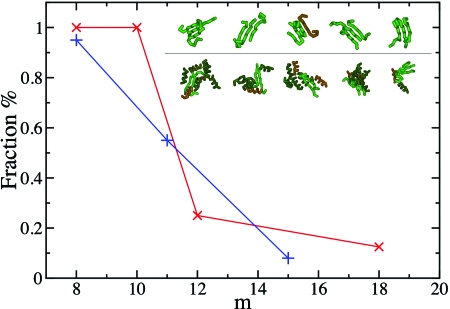
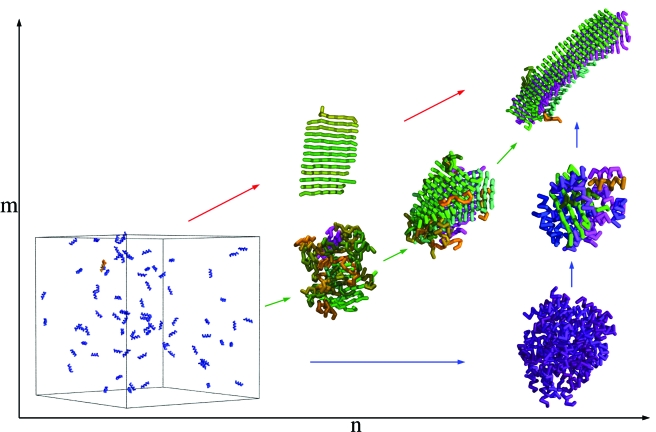
Despite the complexity and the specificity of the amino acid code, a variety of peptides and proteins unrelated in sequence and function exhibit a common behavior and assemble into highly organized amyloid fibrils. The formation of such aggregates is often described by a nucleation and growth mechanism, in which the proteins involved also form intermediate oligomeric aggregates before they reorganize and grow into ordered fibrils with a characteristic cross-beta structure. It is extremely difficult to experimentally obtain an accurate description of the early stages of this phenomenon due to the transient nature and structural heterogeneity of the oligomeric precursors. We investigate here the phenomenon of ordered aggregation by using the recently introduced tube model of polypeptide chains in conjunction with the generic hypothesis of amyloid formation. Under conditions where oligomer formation is a rare event-the most common conditions for forming amyloid fibrils by experiment-we calculate directly the nucleation barriers associated with oligomer formation and conversion into cross-beta structure in order to reveal the nature of these species, determine the critical nuclei, and characterize their dependence on the hydrophobicity of the peptides and the thermodynamic parameters associated with aggregation and amyloid formation.
Figures
References
-
- Bader, R, Bamford, R, Zurdo, J, Luisi, B F, and Dobson, C M (2006). “Probing the mechanism of amyloidogenesis through a tandem repeat of the PI3-SH3 domain suggests a generic model for protein aggregation and fibril formation.” J. Mol. Biol. JMOBAK10.1016/j.jmb.2005.11.034 356, 189–208. - DOI - PubMed
-
- Ban, T, Hoshino, M, Takahashi, S, Hamada, D, Hasegawa, K, Naiki, H, and Goto, Y (2004). “Direct observation of A beta amyloid fibril growth and inhibition.” J. Mol. Biol. JMOBAK 344, 757–767. - PubMed
-
- Bieschke, J, Zhang, Q H, Powers, E T, Lerner, R A, and Kelly, J W (2005). “Oxidative metabolites accelerate Alzheimer’s amyloidogenesis by a two-step mechanism, eliminating the requirement for nucleation.” Biochemistry BICHAW 44, 4977–4983. - PubMed
LinkOut - more resources
Full Text Sources