

Collective behavior in animal groups: theoretical models and empirical studies
- PMID: 19404431
- PMCID: PMC2639936
- DOI: 10.2976/1.2961038
Collective behavior in animal groups: theoretical models and empirical studies
Abstract
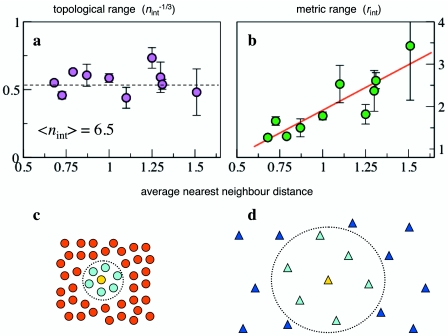
Collective phenomena in animal groups have attracted much attention in the last years, becoming one of the hottest topics in ethology. There are various reasons for this. On the one hand, animal grouping provides a paradigmatic example of self-organization, where collective behavior emerges in absence of centralized control. The mechanism of group formation, where local rules for the individuals lead to a coherent global state, is very general and transcends the detailed nature of its components. In this respect, collective animal behavior is a subject of great interdisciplinary interest. On the other hand, there are several important issues related to the biological function of grouping and its evolutionary success. Research in this field boasts a number of theoretical models, but much less empirical results to compare with. For this reason, even if the general mechanisms through which self-organization is achieved are qualitatively well understood, a quantitative test of the models assumptions is still lacking. New analysis on large groups, which require sophisticated technological procedures, can provide the necessary empirical data.
Figures
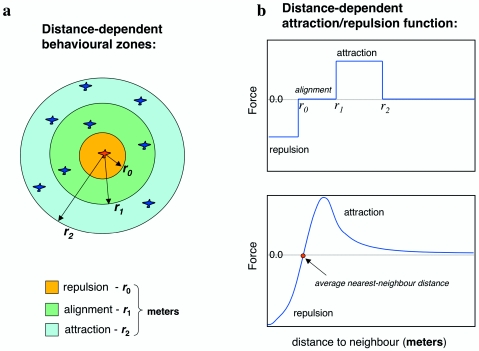
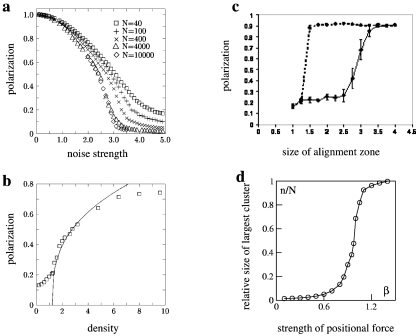
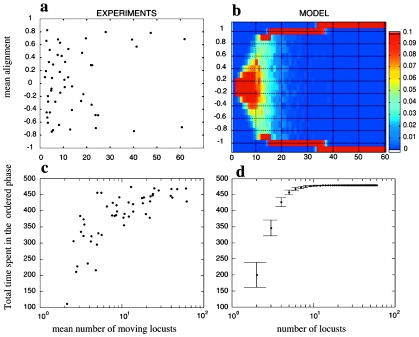
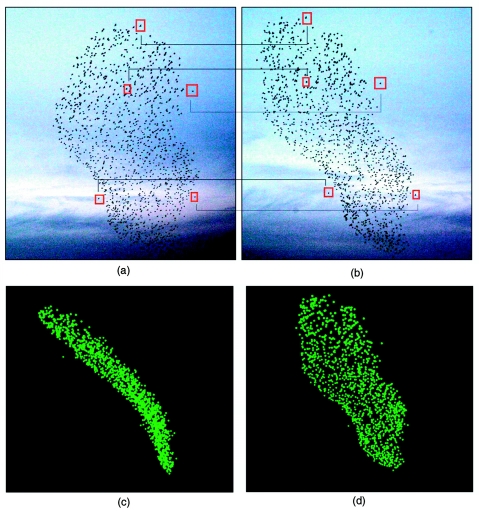
References
-
- Allen, M P, and Tildesley, D J (1989). Computer Simulation of Liquids, Oxford University Press, Oxford.
-
- Aoki, I (1980). “An analysis of the schooling behavior of fish: internal organization and communication process.” Bull. Ocean. Res. Inst. Univ. Tokyo12, 1–65.
-
- Aoki, I (1982). “A simulation study on the schooling mechanism in fish.” Bull. Jpn. Soc. Sci. Fish.48, 1081–1088.
-
- Ballerini, M, Cabibbo, N, Candelier, R, Cavagna, A, Cisbani, E, Giardina, I, Lecomte, V, Orlandi, A, Parisi, G, Procaccini, A, Viale, M, and Zdravkovic, V (2008a). “Interaction ruling animal collective behaviour depends on topological rather than metric distance: evidence from a field study.” Proc. Natl. Acad. Sci. U.S.A. PNASA610.1073/pnas.0711437105 105, 1232–1237. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources