

How molecular motors shape the flagellar beat
- PMID: 19404446
- PMCID: PMC2640991
- DOI: 10.2976/1.2773861
How molecular motors shape the flagellar beat
Abstract
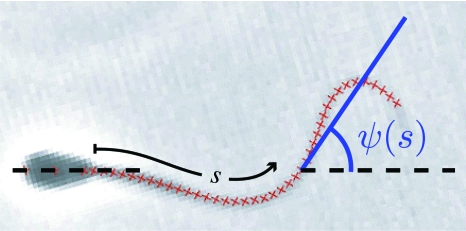
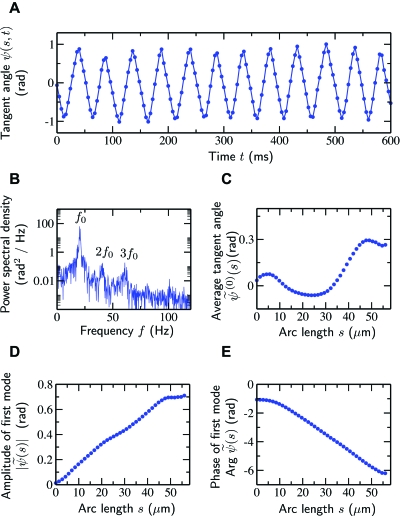
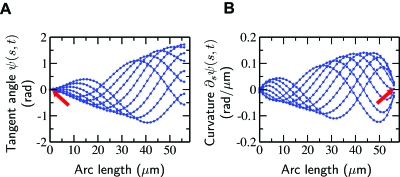
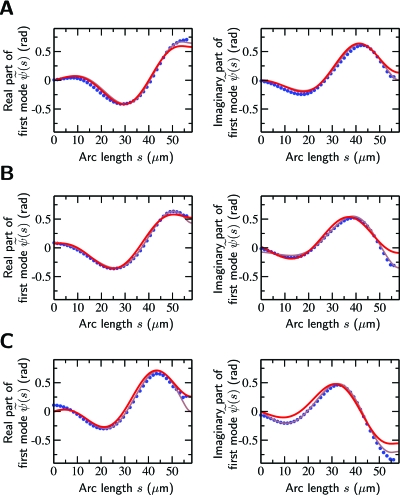
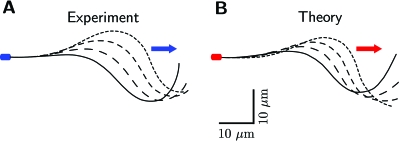
Cilia and eukaryotic flagella are slender cellular appendages whose regular beating propels cells and microorganisms through aqueous media. The beat is an oscillating pattern of propagating bends generated by dynein motor proteins. A key open question is how the activity of the motors is coordinated in space and time. To elucidate the nature of this coordination we inferred the mechanical properties of the motors by analyzing the shape of beating sperm: Steadily beating bull sperm were imaged and their shapes were measured with high precision using a Fourier averaging technique. Comparing our experimental data with wave forms calculated for different scenarios of motor coordination we found that only the scenario of interdoublet sliding regulating motor activity gives rise to satisfactory fits. We propose that the microscopic origin of such "sliding control" is the load dependent detachment rate of motors. Agreement between observed and calculated wave forms was obtained only if significant sliding between microtubules occurred at the base. This suggests a novel mechanism by which changes in basal compliance could reverse the direction of beat propagation. We conclude that the flagellar beat patterns are determined by an interplay of the basal properties of the axoneme and the mechanical feedback of dynein motors.
Figures

References
-
- Afzelius, B A, Dallai, R, Lanzavecchia, S, and Bellon, P L (1995). “Flagellar structure in normal human spermatozoa and in spermatozoa that lack dynein arms.” Tissue Cell 27(3), 241–247. - PubMed
-
- Bray, D (2001). Cell Movements: From Molecules to Motility, 2nd ed., Garland, New York.
-
- Brennen, C, and Winet, H (1977). “Fluid mechanics of propulsion by cilia and flagella.” Annu. Rev. Fluid Mech. 10.1146/annurev.fl.09.010177.002011 9(1), 339–398. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources