

Coupling between neuronal activity and microcirculation: implications for functional brain imaging
- PMID: 19404475
- PMCID: PMC2645573
- DOI: 10.2976/1.2889618
Coupling between neuronal activity and microcirculation: implications for functional brain imaging
Abstract
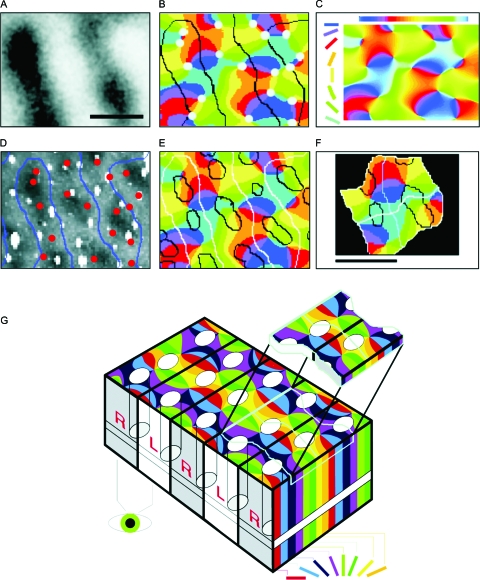
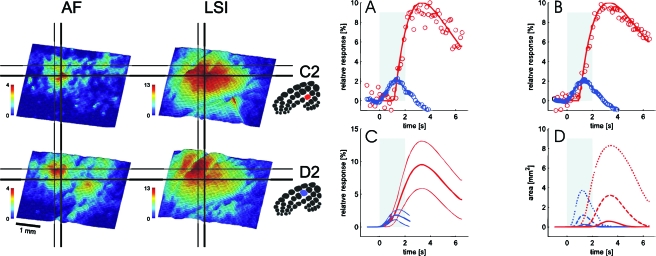
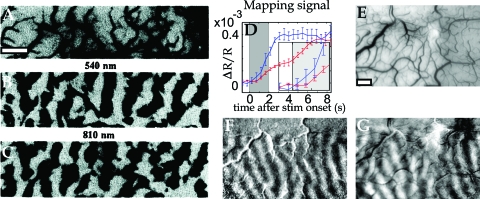
In the neocortex, neurons with similar response properties are often clustered together in column-like structures, giving rise to what has become known as functional architecture-the mapping of various stimulus feature dimensions onto the cortical sheet. At least partially, we owe this finding to the availability of several functional brain imaging techniques, both post-mortem and in-vivo, which have become available over the last two generations, revolutionizing neuroscience by yielding information about the spatial organization of active neurons in the brain. Here, we focus on how our understanding of such functional architecture is linked to the development of those functional imaging methodologies, especially to those that image neuronal activity indirectly, through metabolic or haemodynamic signals, rather than directly through measurement of electrical activity. Some of those approaches allow exploring functional architecture at higher spatial resolution than others. In particular, optical imaging of intrinsic signals reaches the striking detail of approximately 50 mum, and, together with other methodologies, it has allowed characterizing the metabolic and haemodynamic responses induced by sensory-evoked neuronal activity. Here, we review those findings about the spatio-temporal characteristics of neurovascular coupling and discuss their implications for functional brain imaging, including position emission tomography, and non-invasive neuroimaging techniques, such as funtional magnetic resonance imaging, applicable also to the human brain.
Figures
References
-
- Abeles, M (1991). Corticonics: neural circuits of the cerebral cortex, 1st ed., Cambridge University Press, Cambridge.
-
- Akgoren, N, and Lauritzen, M (1999). “Functional recruitment of red blood cells to rat brain microcirculation accompanying increased neuronal activity in cerebellar cortex.” NeuroReport 10, 3257–3263. - PubMed
-
- Altman, D I, Lich, L L, and Powers, W J (1991). “Brief inhalation method to measure cerebral oxygen extraction fraction with PET: accuracy determination under pathologic conditions.” J. Nucl. Med. 32, 1738–1741. - PubMed
LinkOut - more resources
Full Text Sources