

Prostaglandin E2 stimulates the production of amyloid-beta peptides through internalization of the EP4 receptor
- PMID: 19407341
- PMCID: PMC2709369
- DOI: 10.1074/jbc.M109.003269
Prostaglandin E2 stimulates the production of amyloid-beta peptides through internalization of the EP4 receptor
Abstract
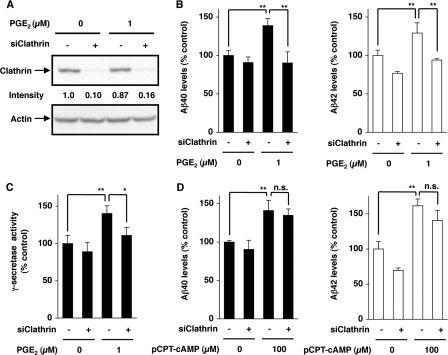
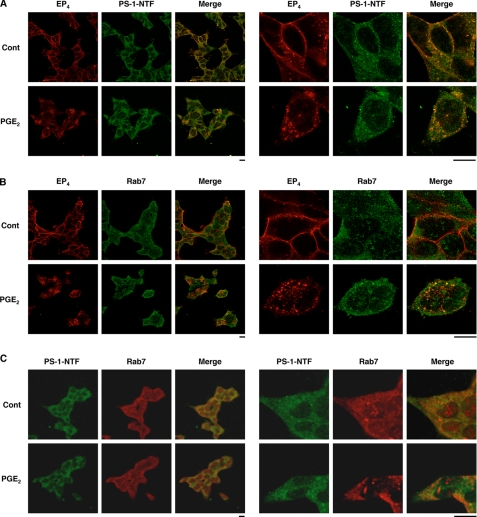
Amyloid-beta (Abeta) peptides, generated by the proteolysis of beta-amyloid precursor protein by beta- and gamma-secretases, play an important role in the pathogenesis of Alzheimer disease. Inflammation is also important. We recently reported that prostaglandin E(2) (PGE(2)), a strong inducer of inflammation, stimulates the production of Abeta through EP(2) and EP(4) receptors, and here we have examined the molecular mechanism. Activation of EP(2) and EP(4) receptors is coupled to an increase in cellular cAMP levels and activation of protein kinase A (PKA). We found that inhibitors of adenylate cyclase and PKA suppress EP(2), but not EP(4), receptor-mediated stimulation of the Abeta production. In contrast, inhibitors of endocytosis suppressed EP(4), but not EP(2), receptor-mediated stimulation. Activation of gamma-secretase was observed with the activation of EP(4) receptors but not EP(2) receptors. PGE(2)-dependent internalization of the EP(4) receptor was observed, and cells expressing a mutant EP(4) receptor lacking the internalization activity did not exhibit PGE(2)-stimulated production of Abeta. A physical interaction between the EP(4) receptor and PS-1, a catalytic subunit of gamma-secretases, was revealed by immunoprecipitation assays. PGE(2)-induced internalization of PS-1 and co-localization of EP(4), PS-1, and Rab7 (a marker of late endosomes and lysosomes) was observed. Co-localization of PS-1 and Rab7 was also observed in the brain of wild-type mice but not of EP(4) receptor null mice. These results suggest that PGE(2)-stimulated production of Abeta involves EP(4) receptor-mediated endocytosis of PS-1 followed by activation of the gamma-secretase, as well as EP(2) receptor-dependent activation of adenylate cyclase and PKA, both of which are important in the inflammation-mediated progression of Alzheimer disease.
Figures





Similar articles
-
Involvement of prostaglandin E2 in production of amyloid-beta peptides both in vitro and in vivo.J Biol Chem. 2007 Nov 9;282(45):32676-88. doi: 10.1074/jbc.M703087200. Epub 2007 Sep 2. J Biol Chem. 2007. PMID: 17767011
-
Improvement of cognitive function in Alzheimer's disease model mice by genetic and pharmacological inhibition of the EP(4) receptor.J Neurochem. 2012 Mar;120(5):795-805. doi: 10.1111/j.1471-4159.2011.07567.x. Epub 2012 Jan 23. J Neurochem. 2012. PMID: 22044482
-
Phosphorylation of glycogen synthase kinase-3 and stimulation of T-cell factor signaling following activation of EP2 and EP4 prostanoid receptors by prostaglandin E2.J Biol Chem. 2002 Jan 25;277(4):2614-9. doi: 10.1074/jbc.M109440200. Epub 2001 Nov 12. J Biol Chem. 2002. PMID: 11706038
-
Roles of the prostaglandin E2 receptors EP subtypes in Alzheimer's disease.Neurosci Bull. 2010 Feb;26(1):77-84. doi: 10.1007/s12264-010-0703-z. Neurosci Bull. 2010. PMID: 20101275 Free PMC article. Review.
-
Prostaglandin E2 and T cells: friends or foes?Immunol Cell Biol. 2012 Jul;90(6):579-86. doi: 10.1038/icb.2011.75. Epub 2011 Sep 27. Immunol Cell Biol. 2012. PMID: 21946663 Free PMC article. Review.
Cited by
-
Therapeutic implications of the prostaglandin pathway in Alzheimer's disease.Biochem Pharmacol. 2014 Apr 15;88(4):565-72. doi: 10.1016/j.bcp.2013.12.014. Epub 2014 Jan 13. Biochem Pharmacol. 2014. PMID: 24434190 Free PMC article. Review.
-
Prostacyclin Promotes Degenerative Pathology in a Model of Alzheimer's Disease.Front Cell Neurosci. 2022 Feb 7;16:769347. doi: 10.3389/fncel.2022.769347. eCollection 2022. Front Cell Neurosci. 2022. PMID: 35197825 Free PMC article.
-
Prostaglandin I2 upregulates the expression of anterior pharynx-defective-1α and anterior pharynx-defective-1β in amyloid precursor protein/presenilin 1 transgenic mice.Aging Cell. 2016 Oct;15(5):861-71. doi: 10.1111/acel.12495. Epub 2016 May 30. Aging Cell. 2016. PMID: 27240539 Free PMC article.
-
Role of prostaglandins in neuroinflammatory and neurodegenerative diseases.Mediators Inflamm. 2012;2012:946813. doi: 10.1155/2012/946813. Epub 2012 Jun 18. Mediators Inflamm. 2012. PMID: 22778499 Free PMC article. Review.
-
Non-Intrinsic, Systemic Mechanisms of Cellular Senescence.Cells. 2023 Dec 5;12(24):2769. doi: 10.3390/cells12242769. Cells. 2023. PMID: 38132089 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
