

Screen for DNA-damage-responsive histone modifications identifies H3K9Ac and H3K56Ac in human cells
- PMID: 19407812
- PMCID: PMC2684025
- DOI: 10.1038/emboj.2009.119
Screen for DNA-damage-responsive histone modifications identifies H3K9Ac and H3K56Ac in human cells
Abstract
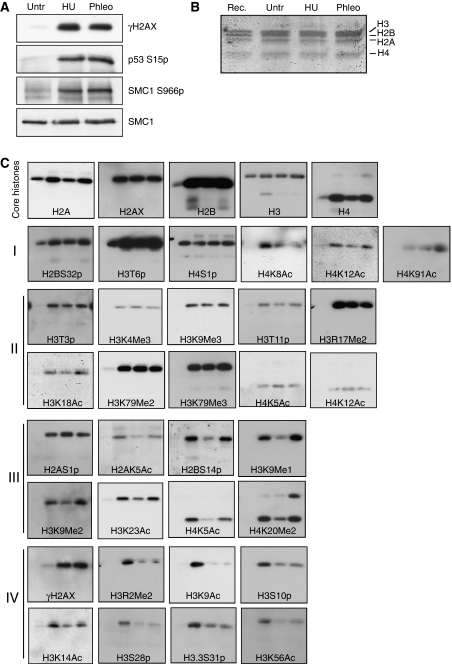
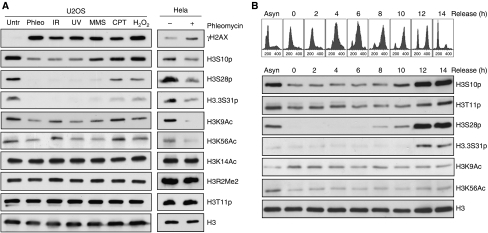
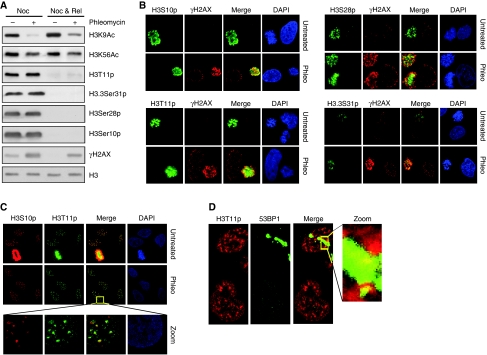
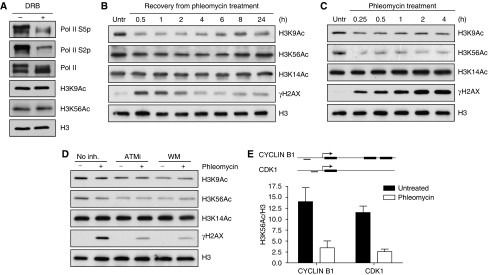
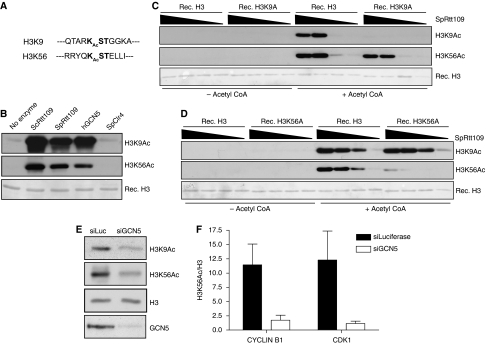
Recognition and repair of damaged DNA occurs within the context of chromatin. The key protein components of chromatin are histones, whose post-translational modifications control diverse chromatin functions. Here, we report our findings from a large-scale screen for DNA-damage-responsive histone modifications in human cells. We have identified specific phosphorylations and acetylations on histone H3 that decrease in response to DNA damage. Significantly, we find that DNA-damage-induced changes in H3S10p, H3S28p and H3.3S31p are a consequence of cell-cycle re-positioning rather than DNA damage per se. In contrast, H3K9Ac and H3K56Ac, a mark previously uncharacterized in human cells, are rapidly and reversibly reduced in response to DNA damage. Finally, we show that the histone acetyl-transferase GCN5/KAT2A acetylates H3K56 in vitro and in vivo. Collectively, our data indicate that though most histone modifications do not change appreciably after genotoxic stress, H3K9Ac and H3K56Ac are reduced in response to DNA damage in human cells.
Figures
Comment in
-
A histone code for the DNA damage response in mammalian cells?EMBO J. 2009 Jul 8;28(13):1828-30. doi: 10.1038/emboj.2009.180. EMBO J. 2009. PMID: 19587679 Free PMC article. No abstract available.
Similar articles
-
Damaged DNA-binding protein down-regulates epigenetic mark H3K56Ac through histone deacetylase 1 and 2.Mutat Res. 2015 Jun;776:16-23. doi: 10.1016/j.mrfmmm.2015.01.005. Epub 2015 Jan 24. Mutat Res. 2015. PMID: 26255936 Free PMC article.
-
Dynamic alteration in H3 serine 10 phosphorylation is G1-phase specific during ionization radiation induced DNA damage response in human cells.Mutat Res. 2015 Mar;773:83-91. doi: 10.1016/j.mrfmmm.2015.01.017. Epub 2015 Feb 9. Mutat Res. 2015. PMID: 25847424
-
Structural insight into the recognition of acetylated histone H3K56ac mediated by the bromodomain of CREB-binding protein.FEBS J. 2017 Oct;284(20):3422-3436. doi: 10.1111/febs.14198. Epub 2017 Sep 12. FEBS J. 2017. PMID: 28815970
-
Role of histone acetyltransferases MOF and Tip60 in genome stability.DNA Repair (Amst). 2021 Nov;107:103205. doi: 10.1016/j.dnarep.2021.103205. Epub 2021 Aug 8. DNA Repair (Amst). 2021. PMID: 34399315 Review.
-
Mammalian DNA repair: HATs and HDACs make their mark through histone acetylation.Mutat Res. 2013 Oct;750(1-2):23-30. doi: 10.1016/j.mrfmmm.2013.07.002. Epub 2013 Aug 6. Mutat Res. 2013. PMID: 23927873 Review.
Cited by
-
Emerging roles for histone modifications in DNA excision repair.FEMS Yeast Res. 2016 Nov;16(7):fow090. doi: 10.1093/femsyr/fow090. Epub 2016 Oct 12. FEMS Yeast Res. 2016. PMID: 27737893 Free PMC article. Review.
-
Kdm4b histone demethylase is a DNA damage response protein and confers a survival advantage following γ-irradiation.J Biol Chem. 2013 Jul 19;288(29):21376-21388. doi: 10.1074/jbc.M113.491514. Epub 2013 Jun 6. J Biol Chem. 2013. PMID: 23744078 Free PMC article.
-
Functions of Ubiquitin and SUMO in DNA Replication and Replication Stress.Front Genet. 2016 May 13;7:87. doi: 10.3389/fgene.2016.00087. eCollection 2016. Front Genet. 2016. PMID: 27242895 Free PMC article. Review.
-
H3 Acetylation-Induced Basal Progenitor Generation and Neocortex Expansion Depends on the Transcription Factor Pax6.Biology (Basel). 2024 Jan 23;13(2):68. doi: 10.3390/biology13020068. Biology (Basel). 2024. PMID: 38392287 Free PMC article.
-
The genetic and epigenetic journey of embryonic stem cells into mature neural cells.Front Genet. 2012 May 18;3:81. doi: 10.3389/fgene.2012.00081. eCollection 2012. Front Genet. 2012. PMID: 22629283 Free PMC article.
References
-
- Aguilera A, Gomez-Gonzalez B (2008) Genome instability: a mechanistic view of its causes and consequences. Nat Rev Genet 9: 204–217 - PubMed
-
- Allison SJ, Milner J (2003) Loss of p53 has site-specific effects on histone H3 modification, including serine 10 phosphorylation important for maintenance of ploidy. Cancer Res 63: 6674–6679 - PubMed
-
- Berger SL (2007) The complex language of chromatin regulation during transcription. Nature 447: 407–412 - PubMed
-
- Camphausen K, Tofilon PJ (2007) Inhibition of histone deacetylation: a strategy for tumor radiosensitization. J Clin Oncol 25: 4051–4056 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
