

MAPL is a new mitochondrial SUMO E3 ligase that regulates mitochondrial fission
- PMID: 19407830
- PMCID: PMC2727426
- DOI: 10.1038/embor.2009.86
MAPL is a new mitochondrial SUMO E3 ligase that regulates mitochondrial fission
Abstract
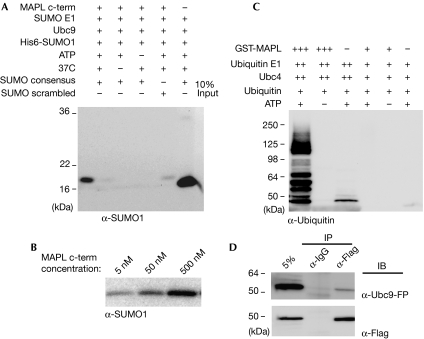
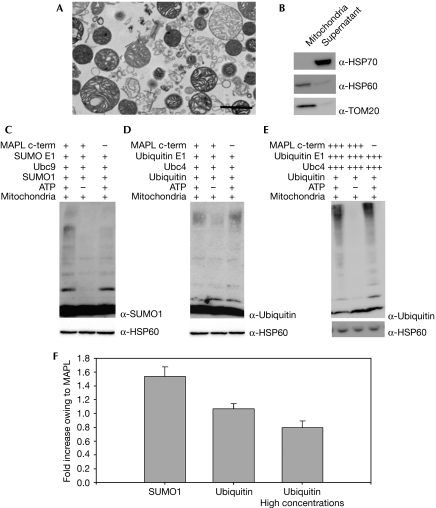
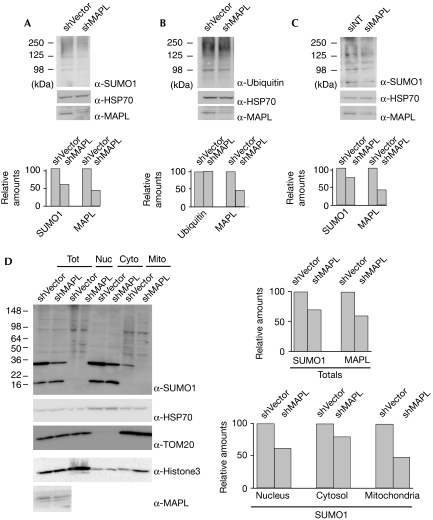
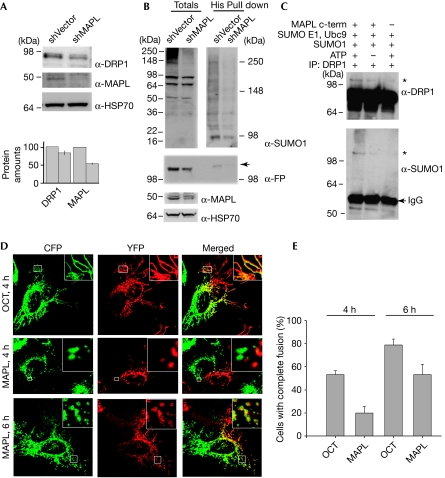
The modification of proteins by the small ubiquitin-like modifier (SUMO) is known to regulate an increasing array of cellular processes. SUMOylation of the mitochondrial fission GTPase dynamin-related protein 1 (DRP1) stimulates mitochondrial fission, suggesting that SUMOylation has an important function in mitochondrial dynamics. The conjugation of SUMO to its substrates requires a regulatory SUMO E3 ligase; however, so far, none has been functionally associated with the mitochondria. By using biochemical assays, overexpression and RNA interference experiments, we characterized the mitochondrial-anchored protein ligase (MAPL) as the first mitochondrial-anchored SUMO E3 ligase. Furthermore, we show that DRP1 is a substrate for MAPL, providing a direct link between MAPL and the fission machinery. Importantly, the large number of unidentified mitochondrial SUMO targets suggests a global role for SUMOylation in mitochondrial function, placing MAPL as a crucial component in the regulation of multiple conjugation events.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Bischof O, Schwamborn K, Martin N, Werner A, Sustmann C, Grosschedl R, Dejean A (2006) The E3 SUMO ligase PIASy is a regulator of cellular senescence and apoptosis. Mol Cell 22: 783–794 - PubMed
-
- Dadke S et al. (2007) Regulation of protein tyrosine phosphatase 1B by sumoylation. Nat Cell Biol 9: 80–85 - PubMed
-
- Geiss-Friedlander R, Melchior F (2007) Concepts in sumoylation: a decade on. Nat Rev Mol Cell Biol 8: 947–956 - PubMed
-
- Harder Z, Zunino R, McBride H (2004) Sumo1 conjugates mitochondrial substrates and participates in mitochondrial fission. Curr Biol 14: 340–345 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
