

The myosin C-loop is an allosteric actin contact sensor in actomyosin
- PMID: 19408946
- PMCID: PMC2759872
- DOI: 10.1021/bi900584q
The myosin C-loop is an allosteric actin contact sensor in actomyosin
Abstract
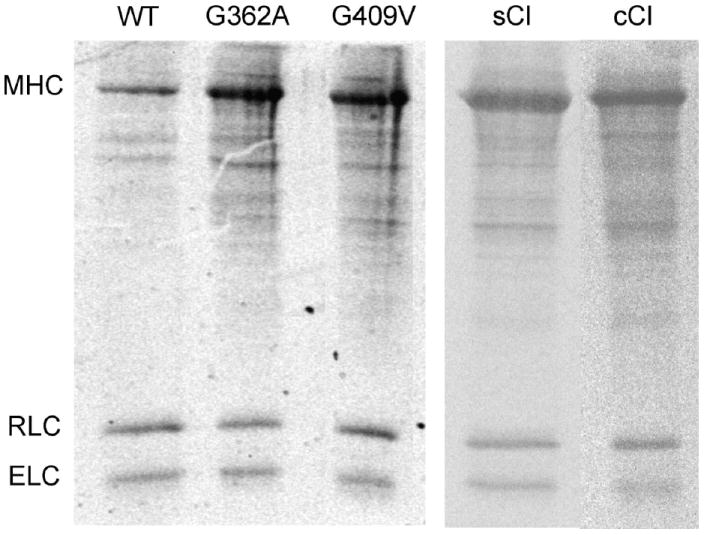
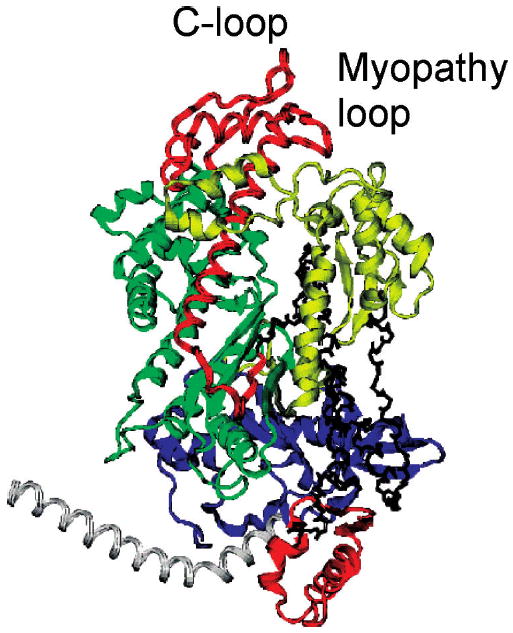
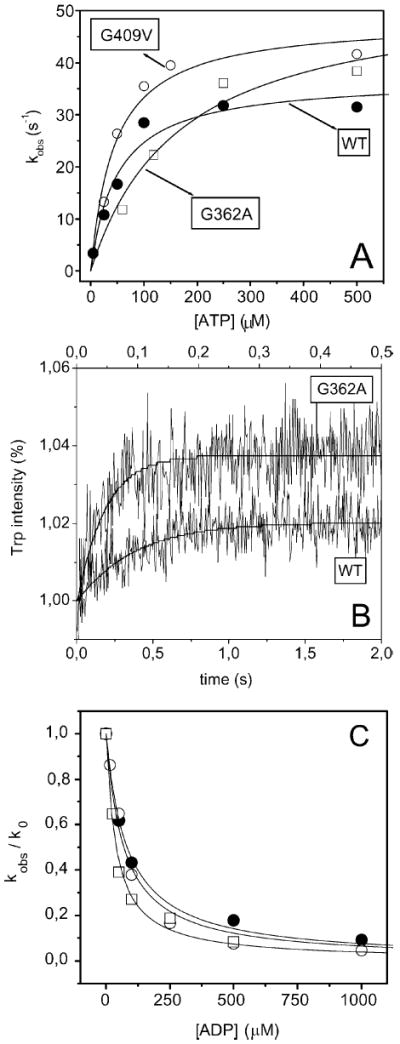
Actin and myosin form the molecular motor in muscle. Myosin is the enzyme performing ATP hydrolysis under the allosteric control of actin such that actin binding initiates product release and force generation in the myosin power stroke. Non-equilibrium Monte Carlo molecular dynamics simulation of the power stroke suggested that a structured surface loop on myosin, the C-loop, is the actin contact sensor initiating actin activation of the myosin ATPase. Previous experimental work demonstrated C-loop binds actin and established the forward and reverse allosteric link between the C-loop and the myosin active site. Here, smooth muscle heavy meromyosin C-loop chimeras were constructed with skeletal (sCl) and cardiac (cCl) myosin C-loops substituted for the native sequence. In both cases, actin-activated ATPase inhibition is indicated mainly by the lower V(max). In vitro motility was also inhibited in the chimeras. Motility data were collected as a function of myosin surface density, with unregulated actin, and with skeletal and cardiac isoforms of tropomyosin-bound actin for the wild type, cCl, and sCl. Slow and fast subpopulations of myosin velocities in the wild-type species were discovered and represent geometrically unfavorable and favorable actomyosin interactions, respectively. Unfavorable interactions are detected at all surface densities tested. Favorable interactions are more probable at higher myosin surface densities. Cardiac tropomyosin-bound actin promotes the favorable actomyosin interactions by lowering the inhibiting geometrical constraint barriers with a structural effect on actin. Neither higher surface density nor cardiac tropomyosin-bound actin can accelerate motility velocity in cCl or sCl, suggesting the element initiating maximal myosin activation by actin resides in the C-loop.
Figures


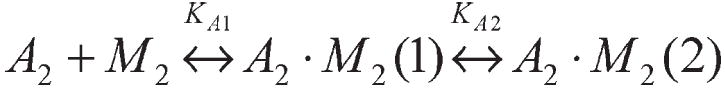



Similar articles
-
Functional characterization of the secondary actin binding site of myosin II.Biochemistry. 1999 Nov 16;38(46):15078-85. doi: 10.1021/bi991595h. Biochemistry. 1999. PMID: 10563790
-
Myosin surface loop 4 modulates inhibition of actomyosin 1b ATPase activity by tropomyosin.Biochemistry. 2007 Mar 13;46(10):2779-86. doi: 10.1021/bi602439f. Epub 2007 Feb 14. Biochemistry. 2007. PMID: 17298083
-
The myosin cardiac loop participates functionally in the actomyosin interaction.J Biol Chem. 2004 May 28;279(22):23415-21. doi: 10.1074/jbc.M310775200. Epub 2004 Mar 11. J Biol Chem. 2004. PMID: 15020589
-
Review: The ATPase mechanism of myosin and actomyosin.Biopolymers. 2016 Aug;105(8):483-91. doi: 10.1002/bip.22853. Biopolymers. 2016. PMID: 27061920 Review.
-
Actomyosin: law and order in motility.Curr Opin Cell Biol. 2000 Feb;12(1):26-34. doi: 10.1016/s0955-0674(99)00053-8. Curr Opin Cell Biol. 2000. PMID: 10679363 Review.
Cited by
-
The R369 Myosin Residue within Loop 4 Is Critical for Actin Binding and Muscle Function in Drosophila.Int J Mol Sci. 2022 Feb 25;23(5):2533. doi: 10.3390/ijms23052533. Int J Mol Sci. 2022. PMID: 35269675 Free PMC article.
-
Single myosin cross-bridge orientation in cardiac papillary muscle detects lever-arm shear strain in transduction.Biochemistry. 2011 Sep 13;50(36):7809-21. doi: 10.1021/bi2008992. Epub 2011 Aug 18. Biochemistry. 2011. PMID: 21819137 Free PMC article.
-
Smooth muscle myosin light chain kinase efficiently phosphorylates serine 15 of cardiac myosin regulatory light chain.Biochem Biophys Res Commun. 2011 Dec 16;416(3-4):367-71. doi: 10.1016/j.bbrc.2011.11.044. Epub 2011 Nov 19. Biochem Biophys Res Commun. 2011. PMID: 22120626 Free PMC article.
-
Natural variant frequencies across domains from different sarcomere proteins cross-correlate to identify inter-protein contacts associated with cardiac muscle function and disease.Mol Biomed. 2021 Nov 15;2(1):35. doi: 10.1186/s43556-021-00056-x. Mol Biomed. 2021. PMID: 35006463 Free PMC article.
-
A physical model of cell metabolism.Sci Rep. 2018 May 29;8(1):8349. doi: 10.1038/s41598-018-26724-7. Sci Rep. 2018. PMID: 29844352 Free PMC article.
References
-
- Huxley HE. The mechanism of muscular contraction. Science. 1969;164:1356–1366. - PubMed
-
- Eisenberg E, Hill TL. Muscle contraction and free energy transduction in biological systems. Science. 1985;227:999–1006. - PubMed
-
- Rayment I, Rypniewski WR, Schmidt-Base K, Smith R, Tomchick DR, Benning MM, Winkelmann DA, Wesenberg G, Holden HM. Three-dimensional structure of myosin subfragment-1: A molecular motor. Science. 1993;261:50–58. - PubMed
-
- Dominguez R, Freyzon Y, Trybus KM, Cohen C. Crystal structure of a vertebrate smooth muscle myosin motor domain and its complex with the essential light chain: Visualization of the pre-power stroke state. Cell. 1998;94:559–571. - PubMed
-
- Bálint M, Sréter FA, Wolf I, Nagy B, Gergely J. The substructure of heavy meromyosin. The effect of Ca2+ and Mg2+ on the tryptic fragmentation of heavy meromyosin. J Biol Chem. 1975;250:6168–6177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
