

Sympathetic-correlated c-Fos expression in the neonatal rat spinal cord in vitro
- PMID: 19409080
- PMCID: PMC2687431
- DOI: 10.1186/1423-0127-16-44
Sympathetic-correlated c-Fos expression in the neonatal rat spinal cord in vitro
Abstract
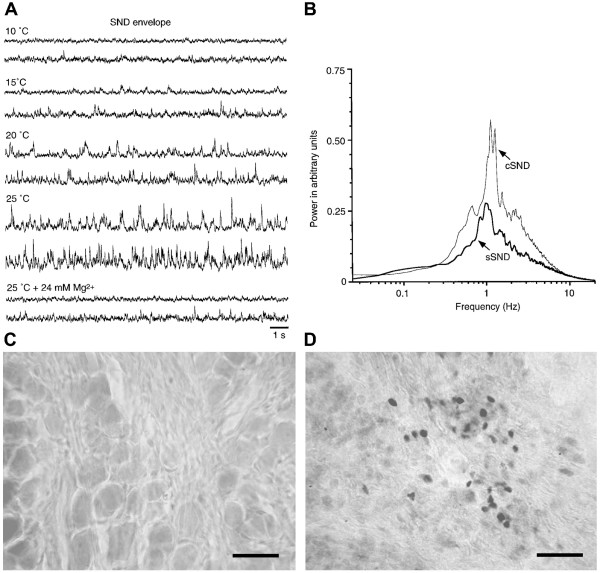
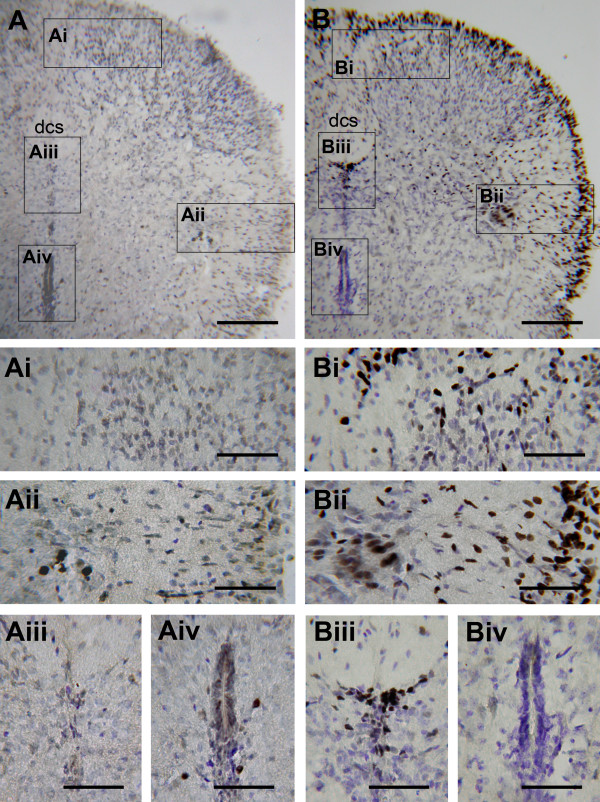
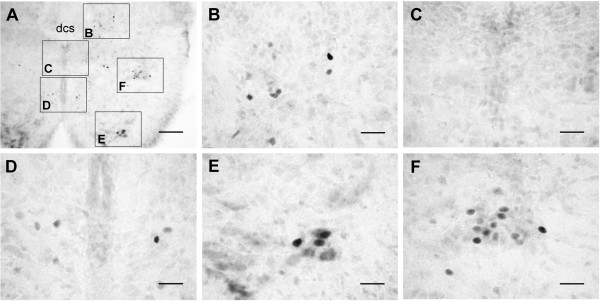
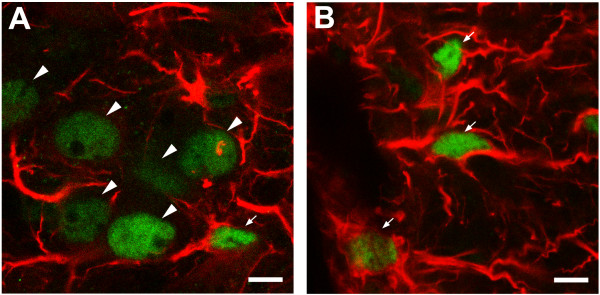
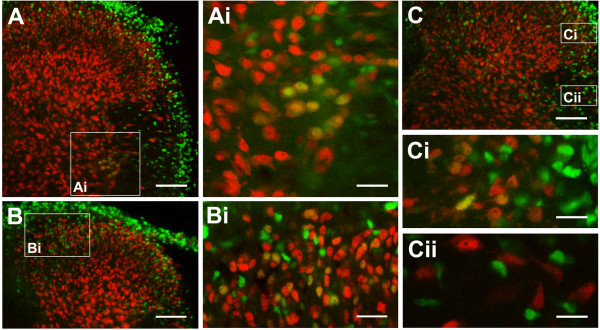
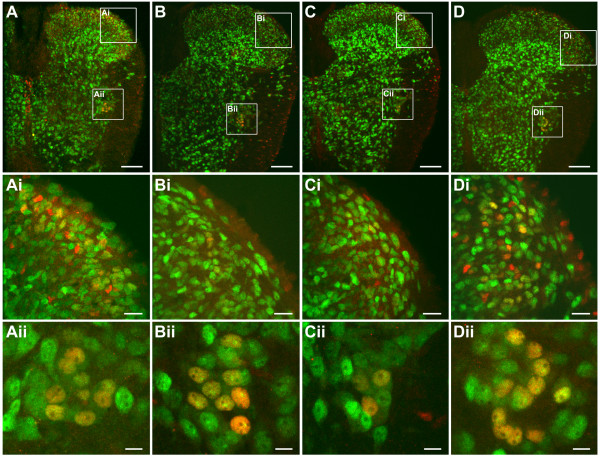
An isolated thoracic spinal cord of the neonatal rat in vitro spontaneously generates sympathetic nerve discharge (SND) at ~25 degrees C, but it fails in SND genesis at < or = 10 degrees C. Basal levels of the c-Fos expression in the spinal cords incubated at < or = 10 degrees C and ~25 degrees C were compared to determine the anatomical substrates that might participate in SND genesis. Cells that exhibited c-Fos immunoreactivity were virtually absent in the spinal cords incubated at < or = 10 degrees C. However, in the spinal cords incubated at ~25 degrees C, c-Fos-positive cells were found in the dorsal laminae, the white matter, lamina X, and the intermediolateral cell column (IML). Cell identities were verified by double labeling of c-Fos with neuron-specific nuclear protein (NeuN), glial fibrillary acidic protein (GFAP), or choline acetyltransferase (ChAT). The c-Fos-positive cells distributed in the white matter and lamina X were NeuN-negative or GFAP-positive and were glial cells. Endogenously active neurons showing c-Fos and NeuN double labeling were scattered in the dorsal laminae and concentrated in the IML. Double labeling of c-Fos and ChAT confirmed the presence of active sympathetic preganglionic neurons (SPNs) in the IML. Suppression of SND genesis by tetrodotoxin (TTX) or mecamylamine (MECA, nicotinic receptor blocker) almost abolished c-Fos expression in dorsal laminae, but only mildly affected c-Fos expression in the SPNs. Therefore, c-Fos expression in some SPNs does not require synaptic activation. Our results suggest that spinal SND genesis is initiated from some spontaneously active SPNs, which are capable of TTX- or MECA-resistant c-Fos expression.
Figures



Similar articles
-
Endogenous activation of nicotinic receptors underlies sympathetic tone generation in neonatal rat spinal cord in vitro.Neuropharmacology. 2006 Dec;51(7-8):1120-8. doi: 10.1016/j.neuropharm.2006.06.015. Epub 2006 Aug 10. Neuropharmacology. 2006. PMID: 16904709
-
GFAP and Fos immunoreactivity in lumbo-sacral spinal cord and medulla oblongata after chronic colonic inflammation in rats.World J Gastroenterol. 2005 Aug 21;11(31):4827-32. doi: 10.3748/wjg.v11.i31.4827. World J Gastroenterol. 2005. PMID: 16097052 Free PMC article.
-
Induction of c-fos gene expression by spinal cord transection in the rat.Brain Res. 1997 Jul 18;763(1):21-9. doi: 10.1016/s0006-8993(97)00356-9. Brain Res. 1997. PMID: 9272824
-
Systemic morphine suppresses noxious stimulus-evoked Fos protein-like immunoreactivity in the rat spinal cord.J Neurosci. 1990 Jan;10(1):323-35. doi: 10.1523/JNEUROSCI.10-01-00323.1990. J Neurosci. 1990. PMID: 1688935 Free PMC article.
-
How sympathetic are your spinal cord circuits?Exp Physiol. 2015 Apr 1;100(4):365-71. doi: 10.1113/EP085031. Exp Physiol. 2015. PMID: 25655449 Review.
Cited by
-
Downregulation of PI3Kcb utilizing adenovirus-mediated transfer of siRNA attenuates bone cancer pain.Int J Clin Exp Pathol. 2014 Oct 15;7(11):8127-35. eCollection 2014. Int J Clin Exp Pathol. 2014. PMID: 25550861 Free PMC article.
-
Regulation of breathing and autonomic outflows by chemoreceptors.Compr Physiol. 2014 Oct;4(4):1511-62. doi: 10.1002/cphy.c140004. Compr Physiol. 2014. PMID: 25428853 Free PMC article. Review.
-
ROS-scavenging hydrogen nanotherapy simultaneously targets inflammation and pain in rheumatoid arthritis.Mater Today Bio. 2025 Jul 10;33:102068. doi: 10.1016/j.mtbio.2025.102068. eCollection 2025 Aug. Mater Today Bio. 2025. PMID: 40688671 Free PMC article.
-
Chondroitinase enhances cortical map plasticity and increases functionally active sprouting axons after brain injury.J Neurotrauma. 2013 Jul 15;30(14):1257-69. doi: 10.1089/neu.2012.2737. Epub 2013 Jul 11. J Neurotrauma. 2013. PMID: 23517225 Free PMC article.
References
-
- Barman SM, Orer HS, Gebber GL. The role of the medullary lateral tegmental field in the generation and baroreceptor reflex control of sympathetic nerve discharge in the cat. Ann N Y Acad Sci. 2001;940:270–85. - PubMed
-
- Lipski J, Lin J, Teo MY, van Wyk M. The network vs. pacemaker theory of the activity of RVL presympathetic neurons – a comparison with another putative pacemaker system. Auton Neurosci. 2002;98:85–89. - PubMed
-
- Dampney RA, Horiuchi J, Tagawa T, Fontes MA, Potts PD, Polson JW. Medullary and supramedullary mechanisms regulating sympathetic vasomotor tone. Acta Physiol Scand. 2003;177(3):209–18. - PubMed
-
- Guyenet PG. The sympathetic control of blood pressure. Nat Rev Neurosci. 2006;7(5):335–46. - PubMed
-
- Su C-K, Phoon S-L, Yen C-T. Identification of active thoracic spinal segments responsible for tonic and bursting sympathetic discharge in neonatal rats. Brain Res. 2003;966:288–99. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
